Medicinski stručnjak članka

Nove publikacije
Jajnik
Posljednji pregledao: 04.07.2025

Svi iLive sadržaji medicinski se pregledavaju ili provjeravaju kako bi se osigurala što je moguće točnija činjenica.
Imamo stroge smjernice za pronalaženje izvora i samo povezujemo s uglednim medijskim stranicama, akademskim istraživačkim institucijama i, kad god je to moguće, medicinski pregledanim studijama. Imajte na umu da su brojevi u zagradama ([1], [2], itd.) Poveznice koje se mogu kliknuti na ove studije.
Ako smatrate da je bilo koji od naših sadržaja netočan, zastario ili na neki drugi način upitan, odaberite ga i pritisnite Ctrl + Enter.
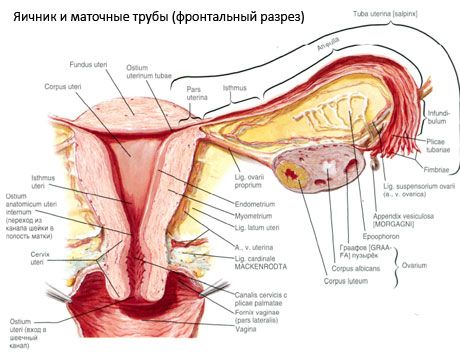
Jajnik (ovarium; grčki oophoron) je parni organ, ženska spolna žlijezda, smještena u zdjeličnoj šupljini iza širokog ligamenta maternice. U jajnicima se razvijaju i sazrijevaju ženske spolne stanice (jaja), te se stvaraju ženski spolni hormoni koji ulaze u krv i limfu. Jajnik ima jajolik oblik, donekle spljošten u anteroposteriornom smjeru. Boja jajnika je ružičasta. Na površini jajnika žene koja je rodila vidljiva su udubljenja i ožiljci - tragovi ovulacije i transformacije žutih tijela. Jajnik teži 5-8 g. Dimenzije jajnika su: duljina 2,5-5,5 cm, širina 1,5-3,0 cm, debljina - do 2 cm. Jajnik ima dvije slobodne površine: medijalnu površinu (facies medialis), okrenutu prema zdjeličnoj šupljini, djelomično prekrivenu jajovodom, i bočnu površinu (facies lateralis), uz bočnu stijenku zdjelice, do blago izraženog udubljenja - jajničke jame. Ova se jama nalazi u kutu između vanjskih ilijakalnih žila prekrivenih peritoneumom na vrhu i arterija maternice i obturatora na dnu. Iza jajnika, ureter odgovarajuće strane prolazi retroperitonealno odozgo prema dolje.
Površine jajnika prelaze u konveksni slobodni (stražnji) rub (margo liber), sprijeda - u mezenterični rub (margo mesovaricus), pričvršćen kratkim naborom peritoneuma (mezenterija jajnika) na stražnji listić širokog ligamenta maternice. Na ovom prednjem rubu organa nalazi se užljebljena udubina - hilum jajnika (hilum ovarii), kroz koji arterija i živci ulaze u jajnik, a vene i limfne žile izlaze. Jajnik također ima dva kraja: zaobljeni gornji tubularni kraj (extremitas tubaria), okrenut prema jajovodu, i donji maternični kraj (extremitas utenna), povezan s maternicom ligamentom jajnika (lig. ovarii proprium). Ovaj ligament, u obliku okruglog užeta debljine oko 6 mm, ide od materničnog kraja jajnika do bočnog kuta maternice, smještenog između dva lista širokog ligamenta maternice. Ligamentozni aparat jajnika također uključuje ligament suspenzorij jajnika (lig.suspensorium ovarii), koji je nabor peritoneuma koji se proteže od stijenke male zdjelice do jajnika i sadrži jajničke žile i snopove vlaknastih vlakana unutra. Jajnik je fiksiran kratkim mezenterijom (mesovarium), koji je duplikat peritoneuma koji se proteže od stražnjeg listića širokog ligamenta maternice do mezenteričnog ruba jajnika. Sami jajnici nisu prekriveni peritoneumom. Najveći jajnički rub jajovoda pričvršćen je na tubularni kraj jajnika. Topografija jajnika ovisi o položaju maternice, njezinoj veličini (tijekom trudnoće). Jajnici su vrlo pokretni organi male zdjelične šupljine.
 [
[ Krvne žile i živci jajnika
Opskrbu krvlju jajnika osiguravaju aa. et vv. ovaricae et uterinae. Obje jajničke arterije (aa. ovaricae dextra et sinistra) potječu s prednje površine aorte odmah ispod bubrežnih arterija; desna obično potječe iz aorte, a lijeva iz bubrežne arterije. Usmjeravajući se prema dolje i lateralno duž prednje površine mišića psoas major, svaka jajnička arterija prelazi sprijeda ureter (dajući mu grane), vanjske ilijačne žile, graničnu liniju i ulazi u zdjeličnu šupljinu, gdje se nalazi u suspenzornom ligamentu jajnika. Slijedeći u medijalnom smjeru, jajnička arterija prolazi između listova širokog ligamenta maternice ispod jajovoda, dajući mu grane, a zatim u mezenterij jajnika; ulazi u hilum jajnika.
Grane jajničke arterije široko anastomoziraju s jajničkim granama maternične arterije. Venski odljev iz jajnika provodi se prvenstveno u jajnički venski pleksus, koji se nalazi u području jajničkog hiluma. Odavde odljev krvi prolazi u dva smjera: kroz maternične i jajničke vene. Desna jajnička vena ima zaliske i ulijeva se u donju šuplju venu. Lijeva jajnička vena ulijeva se u lijevu bubrežnu venu, iako nema zaliske.
Limfna drenaža iz jajnika odvija se kroz limfne žile, posebno obilno u području vrata organa, gdje se razlikuje subovarijalni limfni pleksus. Zatim se limfa ispušta duž limfnih žila jajnika do paraaortalnih limfnih čvorova.
Inervacija jajnika
Simpatički - osiguravaju ga postganglijska vlakna iz celijakijskog (solarnog), gornjeg mezenteričkog i hipogastričnog pleksusa; parasimpatički - visceralni sakralni živci.
Struktura jajnika
Površina jajnika prekrivena je jednoslojnim germinativnim epitelom. Ispod njega leži gusti proteinski omotač od vezivnog tkiva (tunica albuginea). Vezivno tkivo jajnika tvori njegovu stromu (stroma ovarii), bogatu elastičnim vlaknima. Tvar jajnika, njegov parenhim, podijeljena je na vanjski i unutarnji sloj. Unutarnji sloj, smješten u središtu jajnika, bliže njegovim vratima, naziva se srž (medulla ovarii). U tom sloju, u rahlom vezivnom tkivu, nalaze se brojne krvne i limfne žile te živci. Vanjski sloj jajnika - korteks (cortex ovarii) je gušći. Sadrži mnogo vezivnog tkiva, u kojem se nalaze zreli primarni folikuli jajnika (folliculi ovarici primarii), sekundarni (vezikularni) folikuli (folliculi ovarici secundarii, s.vesiculosi), kao i zreli folikuli, Graafovi folikuli (folliculi ovarici maturis), kao i žuta i atretična tjelešca.
Svaki folikul sadrži žensku reproduktivnu jajnu stanicu ili oocitu (ovocit). Jajna stanica je promjera do 150 µm, okrugla je, sadrži jezgru, veliku količinu citoplazme koja, osim staničnih organela, sadrži proteinsko-lipidne inkluzije (žumanjak), glikogen, neophodan za prehranu jajne stanice. Jajna stanica obično potroši svoju zalihu hranjivih tvari unutar 12-24 sata nakon ovulacije. Ako ne dođe do oplodnje, jajna stanica umire.
Ljudska jajna stanica ima dvije membrane koje je prekrivaju. Unutra se nalazi citolema, citoplazmatska membrana jajne stanice. Izvan citoleme nalazi se sloj takozvanih folikularnih stanica koje štite jajnu stanicu i obavljaju funkciju stvaranja hormona - luče estrogene.
Fiziološki položaj maternice, jajovoda i jajnika osiguravaju viseći, fiksirajući i potporni aparati koji ujedinjuju peritoneum, ligamente i tkivo zdjelice. Viseći aparat predstavljen je parnim formacijama, a uključuje okrugle i široke ligamente maternice, vlastite ligamente i viseće ligamente jajnika. Široki ligamenti maternice, vlastiti i viseći ligamenti jajnika drže maternicu u srednjem položaju. Okrugli ligamenti vuku fundus maternice prema naprijed i osiguravaju njegov fiziološki nagib.
Aparat za fiksiranje (sidrenje) osigurava položaj maternice u središtu male zdjelice i praktički onemogućuje njezino pomicanje u strane, naprijed-natrag. No, budući da se ligamentni aparat odvaja od maternice u donjem dijelu, mogući su nagibi maternice u različitim smjerovima. Aparat za fiksiranje uključuje ligamente smještene u labavom tkivu zdjelice i protežu se od donjeg dijela maternice do bočnih, prednjih i stražnjih stijenki zdjelice: sakro-magične, kardinalne, uterovezikalne i vezikopubične ligamente.
Osim mezovarija, razlikuju se sljedeći ligamenti jajnika:
- suspenzorni ligament jajnika, prethodno označen kao infundibulopelvični ligament. To je nabor peritoneuma s krvnim žilama (a. et v. ovarica) te limfnim žilama i živcima jajnika koji prolaze kroz njega, rastegnut između bočne stijenke zdjelice, lumbalne fascije (u području podjele zajedničke ilijačne arterije na vanjsku i unutarnju) i gornjeg (tubarnog) kraja jajnika;
- Pravi ligament jajnika prolazi između listova širokog ligamenta maternice, bliže stražnjem listu, i spaja donji kraj jajnika s bočnim rubom maternice. Pravi ligament jajnika pričvršćen je za maternicu između početka jajovoda i okruglog ligamenta, iza i iznad potonjeg. U debljini ligamenta prolaze rr. ovarii, koji su završna grana maternične arterije;
- Apendikularno-jajni ligament Ligament se proteže od mezenterija slijepog crijeva do desnog jajnika ili širokog ligamenta maternice u obliku nabora peritoneuma. Ligament je nepostojan i opaža se kod 1/2 - 1/3 žena.
Potporni aparat predstavljen je mišićima i fascijom zdjeličnog dna, podijeljenim na donji, srednji i gornji (unutarnji) sloj.
Najmoćniji je gornji (unutarnji) mišićni sloj, predstavljen parnim mišićem koji podiže anus. Sastoji se od mišićnih snopova koji se šire od trtične kosti do zdjeličnih kostiju u tri smjera (pubococcygeus, iliococcygeus i ischiococcygeus). Ovaj sloj mišića naziva se i zdjelična dijafragma.
Srednji sloj mišića nalazi se između simfize, pubične i ishijalne kosti. Srednji sloj mišića - urogenitalna dijafragma - zauzima prednju polovicu zdjeličnog izlaza, kroz koji prolaze uretra i vagina. U prednjem dijelu između njezinih listova nalaze se mišićni snopovi koji tvore vanjski sfinkter uretre, u stražnjem dijelu su mišićni snopovi koji idu u poprečnom smjeru - duboki poprečni mišić međice.
Donji (vanjski) sloj mišića dna zdjelice sastoji se od površinskih mišića, čiji oblik podsjeća na broj 8. To uključuje bulbokavernozni, ishiokavernozni, vanjski analni sfinkter i površinski transverzalni perinealni mišić.
Ontogeneza jajnika
Proces rasta i atrezije folikula započinje u 20. tjednu trudnoće, a do trenutka poroda u jajnicima djevojčice ostaje do 2 milijuna jajnih stanica. Do menarhe njihov se broj smanjuje na 300 tisuća. Tijekom cijelog reproduktivnog razdoblja, najviše 500 folikula dostigne zrelost i ovulira. Početni rast folikula ne ovisi o stimulaciji FSH-om, ograničen je i brzo dolazi do atrezije. Smatra se da su umjesto steroidnih hormona, lokalni autokrini/parakrini peptidi glavni regulator rasta i atrezije primarnih folikula. Smatra se da proces rasta i atrezije folikula ne prekidaju nikakvi fiziološki procesi. Taj se proces nastavlja u svim dobnim skupinama, uključujući intrauterino razdoblje i menopauzu, a prekida ga trudnoća, ovulacija i anovulacija. Mehanizam koji pokreće rast folikula i njihov broj u svakom specifičnom ciklusu još uvijek nije jasan.
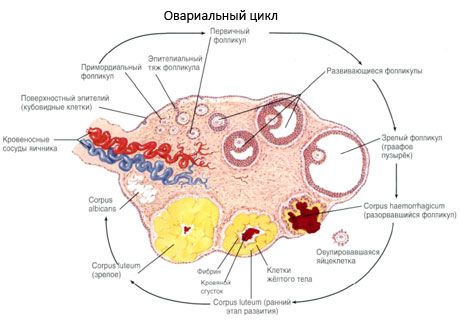
Tijekom svog razvoja, folikul prolazi kroz nekoliko faza razvoja. Primordijalne zametne stanice nastaju iz endoderma žumanjčane vrećice, alantoisa, i migriraju u genitalno područje embrija u 5-6 tjednu trudnoće. Kao rezultat brze mitotičke diobe, koja se nastavlja od 6-8 tjedana do 16-20 tjedna trudnoće, u jajnicima embrija formira se do 6-7 milijuna oocita, okruženih tankim slojem granuloznih stanica.
Preantralni folikul - oocita je okružena membranom (Zona pellucida). Granuloza stanice koje okružuju oocit počinju proliferirati, njihov rast ovisi o gonadotropinima i korelira s razinom estrogena. Granuloza stanice su meta za FSH. U fazi preantralnog folikula, granuloza stanice sposobne su sintetizirati tri klase steroida: pretežno inducira aktivnost aromataze, glavnog enzima koji pretvara androgene u estradiol. Vjeruje se da je estradiol sposoban povećati broj vlastitih receptora, vršeći izravan mitogeni učinak na granuloza stanice neovisno o FSH. Smatra se parakrinim faktorom koji pojačava učinke FSH, uključujući aktivaciju procesa aromatizacije.
FSH receptori pojavljuju se na membranama granuloza stanica čim započne rast folikula. Smanjenje ili povećanje FSH dovodi do promjene broja njegovih receptora. Ovo djelovanje FSH moduliraju faktori rasta. FSH djeluje putem G-proteina, sustava adenilat ciklaze, iako je steroidogeneza u folikulu uglavnom regulirana FSH, u ovaj proces uključeno je mnogo faktora: ionski kanali, receptori tirozin kinaze, fosfolipazni sustav sekundarnih glasnika.
Uloga androgena u ranom razvoju folikula je složena. Granuloza stanice imaju androgene receptore. Oni nisu samo supstrat za FSH-induciranu aromatizaciju u estrogene, već mogu i pojačati aromatizaciju pri niskim koncentracijama. Kada se razina androgena poveća, preantralne granuloza stanice preferiraju ne put aromatizacije u estrogene, već jednostavniji androgeni put putem 5α-reduktaze kako bi se pretvorile u androgen koji se ne može pretvoriti u estrogen, čime se inhibira aktivnost aromataze. Ovaj proces također inhibira stvaranje FSH i LH receptora, čime se zaustavlja razvoj folikula.
Procesom aromatizacije, folikul s visokom razinom androgena prolazi kroz procese atrezije. Rast i razvoj folikula ovisi o njegovoj sposobnosti pretvaranja androgena u estrogene.
U prisutnosti FSH, dominantna tvar folikularne tekućine bit će estrogeni. U odsutnosti FSH - androgeni. LH je normalno odsutan u folikularnoj tekućini do sredine ciklusa. Čim se razina LH poveća, mitotička aktivnost granuloznih stanica se smanjuje, pojavljuju se degenerativne promjene i razina androgena u folikulu se povećava. Razina steroida u folikularnoj tekućini je viša nego u plazmi i odražava funkcionalnu aktivnost stanica jajnika: granuloznih i teka stanica. Ako je jedina meta za FSH granulozne stanice, tada LH ima mnogo ciljeva - teka stanice, stromalne i luteinske stanice te granulozne stanice. I granulozne i teka stanice imaju sposobnost steroidogeneze, ali aromatazna aktivnost prevladava u granuloznim stanicama.
Kao odgovor na LH, teka stanice proizvode androgene, koje granuloza stanice zatim pretvaraju u estrogene putem aromatizacije inducirane FSH-om.
Kako folikul raste, teka stanice počinju eksprimirati gene za LH receptore, P450 sec i 3beta-hidroksisteroid dehidrogenazu, faktor rasta sličan inzulinu (IGF-1) sinergistički s LH povećava ekspresiju gena, ali ne stimulira steroidogenezu.
Ovarijalna steroidogeneza uvijek je ovisna o LH. Kako folikul raste, teka stanice eksprimiraju enzim P450c17, koji iz kolesterola stvara androgene. Granuloza stanice nemaju ovaj enzim i ovisne su o teka stanicama koje proizvode estrogene iz androgena. Za razliku od steroidogeneze, folikulogeneza je ovisna o FSH. Kako folikul raste i razina estrogena se povećava, aktivira se mehanizam povratne sprege - inhibira se proizvodnja FSH, što zauzvrat dovodi do smanjenja aktivnosti aromataze folikula i, u konačnici, do folikularne atrezije putem apoptoze (programirane stanične smrti).
Mehanizam povratne sprege estrogena i FSH inhibira razvoj folikula koji su počeli rasti, ali ne i dominantnog folikula. Dominantni folikul sadrži više FSH receptora, koji podržavaju proliferaciju granuloza stanica i aromatizaciju androgena u estrogene. Osim toga, parakrini i autokrini putevi djeluju kao važan koordinator razvoja antralnog folikula.
Autokrini/parakrini regulator sastoji se od peptida (inhibin, aktivin, folistatin) koje sintetiziraju granuloza stanice kao odgovor na FSH i ulaze u folikularnu tekućinu. Inhibin smanjuje lučenje FSH; aktivin stimulira oslobađanje FSH iz hipofize i pojačava djelovanje FSH u jajniku; folistatin potiskuje aktivnost FSH, moguće vezanjem aktivina. Nakon ovulacije i razvoja žutog tijela, inhibin je pod kontrolom LH.
Rast i diferencijacija stanica jajnika pod utjecajem su faktora rasta sličnih inzulinu (IGE). IGF-1 djeluje na granulosa stanice, uzrokujući povećanje cikličkog adenozin monofosfata (cAMP), progesterona, oksitocina, proteoglikana i inhibina.
IGF-1 djeluje na teka stanice, uzrokujući povećanu proizvodnju androgena. Teka stanice pak proizvode faktor tumorske nekroze (TNF) i epidermalni faktor rasta (EGF), koje također regulira FSH.
EGF stimulira proliferaciju granuloza stanica. IGF-2 je glavni faktor rasta u folikularnoj tekućini, a u njoj se nalaze i IGF-1, TNF-a, TNF-3 i EGF.
Čini se da poremećaj parakrine i/ili autokrine regulacije funkcije jajnika igra ulogu u poremećajima ovulacije i razvoju policističnih jajnika.
Kako antralni folikul raste, sadržaj estrogena u folikularnoj tekućini se povećava. Na vrhuncu njihovog povećanja, receptori za LH pojavljuju se na granuloznim stanicama, dolazi do luteinizacije granuloznih stanica i povećava se proizvodnja progesterona. Dakle, u preovulatornom razdoblju, povećanje proizvodnje estrogena uzrokuje pojavu LH receptora, LH pak uzrokuje luteinizaciju granuloznih stanica i proizvodnju progesterona. Povećanje progesterona smanjuje razinu estrogena, što očito uzrokuje drugi vrhunac FSH u sredini ciklusa.
Smatra se da ovulacija nastupa 10-12 sati nakon vrhunca LH i 24-36 sati nakon vrhunca estradiola. Smatra se da LH stimulira redukcijsku diobu oocita, luteinizaciju granulosa stanica i sintezu progesterona i prostaglandina u folikulu.
Progesteron pojačava aktivnost proteolitičkih enzima koji zajedno s prostaglandinom sudjeluju u pucanju stijenke folikula. Vrhunac FSH izazvan progesteronom omogućuje izlazak oocite iz folikula pretvaranjem plazminogena u proteolitički enzim plazmin, te osigurava dovoljan broj LH receptora za normalan razvoj lutealne faze.
Unutar 3 dana nakon ovulacije, granuloza stanice se povećavaju, a u njima se pojavljuju karakteristične vakuole ispunjene pigmentom, luteinom. Teka-lutealne stanice se diferenciraju od teke i strome te postaju dio žutog tijela. Kapilare koje prodiru u žuto tijelo razvijaju se vrlo brzo pod utjecajem faktora angiogeneze, a s poboljšanom vaskularizacijom povećava se proizvodnja progesterona i estrogena. Aktivnost steroidogeneze i životni vijek žutog tijela određeni su razinom LH. Žuto tijelo nije homogena stanična tvorba. Osim 2 vrste žutih stanica, sadrži endotelne stanice, makrofage, fibroblaste itd. Velike lutealne stanice proizvode peptide (relaksin, oksitocin) i aktivnije su u steroidogenezi s većom aktivnošću aromataze i većom sintezom progesterona od malih stanica.
Vrhunac progesterona javlja se 8. dan nakon vrha LH. Primjećuje se da se progesteron i estradiol epizodično izlučuju u lutealnoj fazi u korelaciji s pulsatilnom proizvodnjom LH. Stvaranjem žutog tijela, kontrola proizvodnje inhibina prelazi s FSH na LH. Inhibin se povećava s porastom estradiola prije vrha LH i nastavlja rasti nakon vrha LH, iako razina estrogena opada. Iako inhibin i estradiol izlučuju granuloza stanice, regulirani su različitim putovima. Pad inhibina na kraju lutealne faze doprinosi porastu FSH za sljedeći ciklus.
Žuto tijelo se vrlo brzo smanjuje - 9.-11. dana nakon ovulacije.
Mehanizam degeneracije nije jasan i nije povezan s luteolitičkom ulogom estrogena ili s mehanizmom povezanim s receptorima, kao što je uočeno u endometriju. Postoji još jedno objašnjenje za ulogu estrogena koje proizvodi žuto tijelo. Poznato je da su estrogeni potrebni za sintezu progesteronskih receptora u endometriju. Estrogeni lutealne faze vjerojatno su neophodni za promjene povezane s progesteronom u endometriju nakon ovulacije. Neadekvatan razvoj progesteronskih receptora, kao posljedica neadekvatne razine estrogena, moguće je dodatni mehanizam za neplodnost i rane gubitke trudnoće, još jedan oblik nedostatka lutealne faze. Vjeruje se da se životni vijek žutog tijela utvrđuje u vrijeme ovulacije. I sigurno će se povući ako ga ne podržava humani korionski gonadotropin zbog trudnoće. Dakle, regresija žutog tijela dovodi do smanjenja razine estradiola, progesterona i inhibina. Smanjenje inhibina uklanja njegov supresivni učinak na FSH; Smanjenje estradiola i progesterona omogućuje vrlo brzu obnovu lučenja GnRH i uklanjanje mehanizma povratne sprege iz hipofize. Smanjenje inhibina i estradiola, zajedno s povećanjem GnRH, dovodi do porasta FSH nad LH. Povećanje FSH dovodi do rasta folikula s naknadnim odabirom dominantnog folikula, a novi ciklus počinje ako ne dođe do trudnoće. Steroidni hormoni igraju vodeću ulogu u reproduktivnoj biologiji i općoj fiziologiji. Oni određuju ljudski fenotip, utječu na kardiovaskularni sustav, metabolizam kostiju, kožu, opće blagostanje i igraju ključnu ulogu u trudnoći. Djelovanje steroidnih hormona odražava unutarstanične i genetske mehanizme koji su potrebni za prijenos izvanstaničnog signala u staničnu jezgru kako bi se izazvao fiziološki odgovor.
Estrogeni difuzno prodiru kroz staničnu membranu i vežu se za receptore smještene u staničnoj jezgri. Kompleks receptor-steroid zatim se veže za DNA. U ciljnim stanicama ove interakcije dovode do ekspresije gena, sinteze proteina i specifične funkcije stanica i tkiva.