
Eksperimentalni modeli osteoartritisa
Posljednji pregledao: 23.04.2024

Svi iLive sadržaji medicinski se pregledavaju ili provjeravaju kako bi se osigurala što je moguće točnija činjenica.
Imamo stroge smjernice za pronalaženje izvora i samo povezujemo s uglednim medijskim stranicama, akademskim istraživačkim institucijama i, kad god je to moguće, medicinski pregledanim studijama. Imajte na umu da su brojevi u zagradama ([1], [2], itd.) Poveznice koje se mogu kliknuti na ove studije.
Ako smatrate da je bilo koji od naših sadržaja netočan, zastario ili na neki drugi način upitan, odaberite ga i pritisnite Ctrl + Enter.
Hrskavica je visoko specijalizirano tkivo koje sadrži samo jednu vrstu stanica (hondrociti), karakteriziran nedostatkom krvnih i limfnih žila. Prehrana hrskavice uglavnom se provodi apsorpcijom iz sinovijalne tekućine. Metabolizam hondro-cita regulira se s brojnim topljivim faktorima koje lokalno proizvode hondrociti i okolna tkiva. Funkcija hondrocita također ovisi o sastavu ekstracelularnog medija (napetost kisika, koncentraciju iona, pH, itd.), VCM sastav, interakciju stanica i matriksa, fizičke signale. Glavni zadatak eksperimentalnog modeliranja je stvaranje kultura u ekstracelularnom okruženju bez promjene fenotipa zrele stanice. Drugi zadatak je stvoriti kulture za proučavanje prijevremenog, odgođenog, kratkog ili dugoročnog odziva hondrocita na kemijske i / ili fizičke signale. Studije in vitro također pružaju priliku za proučavanje ponašanja kondrocita kod osteoartritisa. Treći zadatak je razvoj ko-kurativnih sustava, koji omogućuju proučavanje interakcija različitih tkiva u zglobu. Četvrti zadatak je priprema hrskavičnih implantata za naknadnu transplantaciju. I konačno, peti zadatak je proučiti čimbenike rasta, citokine ili terapeutske agense koji su sposobni stimulirati popravak i / ili inhibirati njegovu resorpciju hrskavice.
Tijekom proteklog desetljeća, stvorila različite obrasce zglobne hrskavice kulturama stanica, među njima - monoslojevi, suspendirane kulture, kulture hondronov, dijelova biljaka, sukulture, kulturi besmrtne stanice. Svaka kultura ima svoje prednosti i nedostatke, a svaka je prikladna za proučavanje jednog posebnog aspekta metabolizma kondrocita. Tako su hrskavični eksplanatori izvrstan model za proučavanje prometa matričnih elemenata koji zahtijevaju originalne receptore stanične površine i normalne interakcije stanica matriksa i matriksa. Istodobno, preporuča se provesti ispitivanje naslaga u matrici ili mehanizmi za regulaciju metabolizma hondrocita, na kulturi izoliranih stanica. Jednooblična kultura niske gustoće je neophodna za proučavanje procesa diferencijacije stanica. Kulture suspendirane u prirodnoj ili sintetskoj matrici su model za analizu adaptivnog odgovora kondrocita prema mehaničkom stresu.
Kulture hondrocita
Pri odabiru hrskavice za in vitro studije potrebno je razmotriti nekoliko važnih točaka. Sastav matriksa i metabolička aktivnost hondrocita variraju u različitim zglobovima, a potonji ovisi o dubini kondrocita u tkivu. Ti su podaci dobiveni u nekoliko pokusa u kojima su proučavane izolirane podpopulacije hondrocita iz hrskavičnih zona različitih dubina. Pronađene su brojne morfološke i biokemijske razlike između kultiviranih hondrocita koji se nalaze u površini i dubokim slojevima zglobne hrskavice. Stanice površine sintetiziraju rijetku, osiromašenu matricu proteoglikanskog fibrila, dok dublje stanice proizvode matricu bogatu fibrilima i proteoglikanima. Štoviše, površinske stanice proizvode relativno manje manje agregirane proteoglikane i hialuronsku kiselinu i relativno manje agrekan i keratan sulfat nego duboko locirane hondrocite. Još jedna važna značajka prepoznatljivosti metabolizma hondrocita izoliranih iz hrskavičnih zona različitih dubina je odgovor na egzogeni poticaj. Prema M. Aydelotte i co-autora, junak hondrociti iz površinske zone hrskavice bili su osjetljiviji na IL-1 od stanica duboke zone.
Ponašanje stanica također ovisi o lokaciji tkiva. Hondrociti hrskavice i uši rubove, uzete iz iste životinje koje reagiraju različito na faktore rasta kao što su faktor rasta fibroblasta (FGF) i TGF-beta. FGF je povećao ugradnju timidina, prolina i leucina u hondrocitnu kulturu rebra, ali ne i uho. TGF-P povećava ugradnju timidina u hrskavice hondrociti rebra i uha, ali nije imalo učinka na timidina u hondrocite i prolin uha. Stanice hrskavice dobivene iz područja s najvećim opterećenjem razlikuju se od onih s mjesta s niskim opterećenjem hrskavice. Primjerice zreli hondrociti iz hrskavice zgloba koljena od središnjeg područja ovaca zglobne površine tibijalne kosti nisu obuhvaćeni meniskusa, koje nosi najveći teret in vivo, manje sintetiziranog agrekan, dekorinom ali veća od stanice područja obuhvaćena meniskusa. Autori također naglašavaju važnost korištenja hrskavice iz identičnih zglobnih zona pri ispitivanju sintetičke funkcije zglobova.
Metabolizam kondrocita i njihov odgovor na regulatorne faktore također značajno ovise o dobi donatora, razvoju kostura i stanju zglobova iz kojih se uzimaju stanice. U ljudskim hondrocitima, opaženo je značajno smanjenje s dobi proliferativnog odgovora. Najveće smanjenje zabilježeno je kod donatora u dobi od 40-50 godina i preko 60 godina. Štoviše, težina proliferativnog odgovora na čimbenike rasta (npr. FGF i TGF-beta) smanjuje se tijekom starenja. Pored kvantitativnih promjena u proliferaciji hondrocita, postoje i kvalitativne promjene. Mlade donorske stanice (10-20 godina) bolje reagiraju na faktor rasta izvedenog s trombocitima (PDGF) nego na TGF-beta, dok se suprotno opaža kod odraslih donorskih stanica. Da bi se objasnile promjene u sintetičkoj funkciji kondrocita i njihov odgovor na učinak čimbenika rasta, koriste se nekoliko mehanizama. Među njima je smanjenje broja i afiniteta površinskih celularnih receptora, promjena sinteze i bioaktivnosti faktora rasta i citokina, modifikacija postreceptorskih signala.
Patološko stanje zglobova također mijenja morfologiju i metaboličku aktivnost kondrocita. Tako su J. Kouri i koautori (1996.) identificirali tri podpopulacije hondrocita u hrskavici s osteoartritisom. Hondrociti iz površinskog i gornjeg dijela hrskavice formiraju klastere i sintetiziraju više proteoglikana i kolagena. TGF-beta, i inzulinu-sličan faktor rasta (IGF) može stimulirati sintezu proteoglikana hondrociti i djelomično neutralizaciji djelovanja IL-1 i TNF-a. Hrskavice pogođeno osteoartritisa, a hondrociti izoliraju se iz hrskavice kod pacijenata s osteoartritisom, su osjetljive na stimuliranje TGF-beta nego zdrave hrskavice kondrocitima. Te su razlike najvjerojatnije povezane s fenotipskim promjenama u hondrocitima u gornjim slojevima zglobne hrskavice.
Izolacija pojedinih hondrocita postiže se sekvencijalnim liječenjem proteolitičkim enzimima ECM-a. Nakon oslobađanja iz ECM, izolirane stanice su idealne za proučavanje sinteze de novo matričnih komponenti . Neki autori koriste samo klostridijsku kolagenazu, drugi preinkubiraju hrskavicu s tripsinom, pronazom, DNazom i / ili hijaluronidazom. Broj izoliranih stanica ovisi o korištenim enzimima. Tako, kada je jedan od obrade kolagenazom 1 g tkiva može se dobiti 1,4T0 6 hondrocite, a kada se koristi pronaza, antibiotik i kolagenazu - 4,3-10 6. Prilikom obrade s kolagenazom, agrekanom, proteini, IL-6, IL-8 ostaju u kulturi stanica mnogo više nego kod sekvencijalnog tretmana s različitim enzimima. Postoji nekoliko objašnjenja za te razlike između dviju kultura stanica:
- Stanični receptori oštećena ili depresivne djelovanjem enzima, TGF-beta inhibira sintezu DNA proteoglikana u novo izoliran kondrocitima (dan 1), a DNA i proteoglikanski sintezu kondrocitima uzgojenih u monosloju (7 dana) stimuliranih TGF-beta. Međutim, kako bi se reekventirale ove membranske komponente potrebno je odgovarajuće razdoblje prije početka eksperimenta.
- Egzogene proteaze mogu prekidati interakciju stanica i matriksa, posredovane integrinima. Integrinska obitelj potiče vezanje kondrocita na VKM molekule (Shakibaei M. I sur., 1997). Ovo rupture mogu utjecati na ekspresiju matričnih gena.
- Ostaci matričnih komponenti mogu regulirati sintetsku funkciju kondrocita. Integrini su u stanju prepoznati proizvode degradacije ECM-a, čime igraju važnu ulogu u popravku tkiva nakon izlaganja proteolitičkim enzimima. T. Larsson i koautori (1989) su izvijestili da dodavanje netaknutih ili fragmentiranih protioglikana u staničnu kulturu potiče sintezu proteina i proteoglikana. Međutim, visoka razina hijaluronske kiseline izaziva značajno smanjenje u uključivanju sulfat proteoglikana sinteze hondrociti pilećih embrija kondrocitima zrele svinjske i hondrosarkom štakora stanice. Osim toga, hijaluronska kiselina - inhibitor proteoglikana oslobađanja iz stanica i u prisutnosti IL-lb, TNF-a, FGF, što pokazuje da je prva suzbijanje biološku aktivnost faktora rasta i citokina. Točan mehanizam na kojem djeluje hijaluronska kiselina ostaje nejasan; Poznato je da hondrociti sadrže receptor za hijaluronsku kiselinu, povezanu s aktinskim filamentima citosola. Vezivanje hijaluronske kiseline na njegov receptor stimulira fosforilaciju proteina. Stoga, ovi podaci pokazuju modulaciju metaboličke funkcije hondrocita fragmentiranim ili nativnim molekulama matričnih proteina aktivacijom membranskih receptorskih stanica.
- Brza stimulacija enzimima sinteze proteina matriksa hondrocitima može biti posljedica promjene u obliku hondrocita i / ili reorganizacije citoskeleta.
- Neki citokini (npr., IL-8) i čimbenici rasta (npr. IGF-1, TGF-P) fiksirani su u ECM. Najpoznatiji primjer je vezanje TGF-beta s dekoracijom, što dovodi do smanjenja sposobnosti bivšeg da inducira stanični rast stanica jajnika kod kineskih hrčaka. Podaci koji sadrže ukras uklanjanja hrskavice s godinama, ukazuju na smanjenje bioraspoloživosti TGF-beta u starenju. Faktori rasta i citokini mogu se osloboditi iz ostataka matriksa tijekom kultura, a zatim modulirati funkciju kondrocita.
Jednodijelna kultura kondrocita
Diferencirani fenotip hondrocita prvenstveno je karakteriziran sintezom tipa II kolagena i tkivno-specifičnih proteoglikana, kao i niska razina mitotičke aktivnosti. Postoje dokazi da je dugoročno uzgoj stanica u jednoslojne, a nakon nekoliko ponovljenih prolaza stanica, hondrociti gube sferični oblik, postaju izduženi, fibroblasta poput oblika. S takvim fibroblasta metaplazija sintetski funkcija također modificirana stanica, karakterizirani progresivno smanjenje u sintezi kolagena II, IX i XI i tipovi poboljšana sinteza kolagena I, III i Utipov. Mali neagregirani proteoglikani sintetiziraju se funkcionalnim agrekanom. Sintetzatapin B i L izuzetno je nizak u diferenciranim stanicama, ali se u procesu gubitka diferencijacije povećava. Kolagenaza-1 se izražava u diferenciranim kondrocitima tijekom produženog uzgoja njegova ekspresija je smanjen, dok povećanje proizvodnje inhibitora tkivnog metaloproteaza (TIMP).
Različiti hondrociti ponovno eksprimiraju kolagen diferenciranog fenotipa kada se prenese iz kulture monoslojne u suspendiranu. Proces diferencijacije vjerojatno je povezan s oblikom stanica. Ova imovina redovito koriste istraživači koji proučavaju oštećene presadnice s autolognim hondrocitima. Mali broj stanica dobivenih iz biopsijskog materijala može se razmnožiti u jednoslojnoj kulturi, a zatim ponovno staviti u trodimenzionalnu matricu prije transplantacije. Re-ekspresije određenog fenotipa dedifferentiated kondrocitima migrirale na agaroznom kulturi, može se potaknuti TGF-p-ossein hidroksiapatita kompleksa i askorbinska kiselina.
Kao odgovor na učinak faktora rasta i citokina, hondrociti se mijenjaju tijekom procesa diferencijacije. Stanični odgovor na citokine i faktore rasta razlikuju se između nediferenciranih i diferenciranih kondrocita. IL-1 stimulira proliferaciju fibroblasta, dok je rast ne-diferenciranih hondrocita inhibiran IL-1. IGF-1 stimulira sintezu DNA u izduženim, ali ne spljoštenim hondrocitima. U diferenciranim hondrocitima, stimulirajući učinci IL-ip i TNF-a na produkte procollagenaze su izraženije nego u nediferenciranim.
Uzgoj hondrocita
Uzgoj hondrocita u suspenziji u tekućem mediju ili u prirodnom ili sintetskom trodimenzionalnom matriksu stabilizira fenotip kondukcije. Stanice zadržavaju svoj sferni oblik, sintetiziraju proteine specifične za tkivo. Preporučena je težinska kultura hondrocita za proučavanje formiranja nove pericellularne matrice. Kulture hondrocita u sintetskim ili prirodnim upojnim polimerima koriste se za implantaciju stanica u defekte hrskavice kako bi stimulirale regeneraciju hrskavice tkiva zgloba. Sintetsko ili prirodno okruženje za implantirane stanice mora zadovoljiti niz zahtjeva:
- Implantati trebaju imati poroznu strukturu za adheziju i rast stanica,
- ni sam polimer ni proizvodi degradacije ne bi trebali uzrokovati upalu ili toksične reakcije tijekom in vivo implantacije ,
- nosač za transplantaciju treba biti sposoban vezati se na susjednu hrskavicu ili subhondralnu kost,
- prirodna ili sintetska matrica mora biti u stanju apsorpcije, njezina razgradnja mora biti uravnotežena pomoću regeneracije tkiva,
- Kako bi se olakšalo popravljanje hrskavice, kemijska struktura i matrična arhitektura matrice trebala bi pomoći u održavanju staničnog fenotipa kojeg kodiraju hondrociti i sintezi proteina specifičnih za tkivo,
- tijekom implantacije in vivo potrebno je proučiti mehanička svojstva sintetičke ili prirodne matrice.
Suspenzija kondrocita u tekućoj fazi
Staničnog u plastičnim posudama, u kojima je uzgoj hondrocita se mogu prevenirati njihove zidove obložene s otopinom metil-celuloza, agaroza, hidrogel (poli-2-hidroksietilmetakrilat) i smjesu kolagena-agaroze. Pod tim uvjetima, hondrociti formiraju klastere i sintetiziraju uglavnom agrekan i tkivne specifične kolagene (II, IX, XI tipovi). Obično se pronađu dvije vrste stanica. Stanice koje se nalaze u sredini zadržavaju sferični oblik okružen dobro razvijenim ECM, što potvrđuje histokemijska i ultrastrukturna istraživanja. Na obodnim hondrocitima imaju diskoidne konture, okruženi su rijetkim ECM; Malo je poznato o funkcionalnim karakteristikama takvih stanica.
Može se uzgajati hondrociti na mikro nosiocima podržanim u suspenziji; kao mikronosača dekstranu kuglice (tsitodeks), kolagenske obložene dekstran kuglice (tsitodeks III), besporovye mikrosfere kolagen tip I (tsellagen). Pod ovim uvjetima kulture, kondrociti se pričvršćuju na površinu mikroprocesora, zadržavaju svoj sferični oblik i proizvode materiju sličnu matrici. Štoviše, upotreba kolagena potiče proliferaciju hondrocita i reeksprimiranje normalnog fenotipa. Stoga, uzgoj hondrocita na mikrosferama kolagena može se upotrijebiti za uspostavljanje staničnog fenotipa prije transplantacije.
Drugi način uzgoja suspenzije hondrocita u tekućem mediju je njihova kultivacija u obliku gustih kuglica koja se sastoji od stanica (0,5-1 x 10 b ) dobivene centrifugiranjem. Takvi hondrociti su sposobni za proizvodnju matriksa koji sadrži velike količine proteoglikana, kolagen tipa II-, ali ne i kolagena tipa I, što je potvrđeno i histološke, imunohistokemijske kvantitativne metode.
Suspenzija kondrocita u prirodnom ECM
Hondrociti se mogu kultivirati u suspenziji u trodimenzionalni matriks (mekom agaru, agaroza, poziva gena ili gel spužve, hijaluronska kiselina, fibrin ljepilo, alginat kuglice).
Kulturni agarozni hondrociti zadržavaju svoj normalni fenotip i sintetiziraju kolagen tip II i tkiva specifični agregat-novi agregati. Kada se uzgajaju u agarozi, stanični sintetizirani proteoglikani se oslobađaju u medij kroz 50 dana. Za usporedbu - u jednoslojnoj kulturi, stanična faza je već prvih 5-6 dana uzgoja nadzirana glikozaminoglikanima; kada se uzgaja u mediju nakon sinteze i otpuštanja glikozaminoglikana, intenzivira se vremenski ovisno smanjenje glikozaminoglikana u prvih 8-10 dana. Ipak, ponašanje hondrocita tijekom njihovog uzgoja u agarozi razlikuje se od onog u uvjetima in vivo. U agarozi, veliki broj sintetiziranih Aggregan agregata sadrži manje i manje molekule nego in vivo. TGF-P stimulira sintezu proteoglikana u eksplantatu, ali smanjuje sintezu agrekana u agarozi.
Alginat je linearni polisaharid dobiven od smeđeg algi. U prisutnosti dvovalentnih kationa, poput Ca 2+ iona, ovaj polimer postaje gel. Svaki hondrocitna uhvaćen u alginata, okružen matrice negativno nabijenih polisaharida, pore koje su usporedive s onima hijaline hrskavice. Matrica koja se formira u hondrocite alginatnih kuglica, koji se sastoji od dva segmenta - tanki sloj matriksa povezanog sa stanicom koji odgovara percelularnim i prostornih matrice zglobne hrskavice, a više udaljeni matriks interterritorial ekvivalenta u nativnom tkivu. Na 30. Dana kulture, relativni i apsolutni volumen koji zauzima stanica, a svaki od dva odjela u alginat zrno je gotovo u potpunosti identični onima u rodnom hrskavice. Za skoro 30 dana hondrociti zadržavaju svoj oblik i proizvodnju sferični agrekanske, hidrodinamičke karakteristike koje su slične onima koje agrekanskih molekula u matriksu hrskavice zgloba, a kolagen molekule II, IX i XI tipova. U isto vrijeme, kao i drugim kulturama, suspenzija, alginat perle na površini spljoštene stanice su prisutne koji stvaraju malu količinu kolagena tipa molekula, oslobađa izravno u okoliš, a ne ugrađen u videorekorder. U zrncima alginata opaža se umjerena proliferacija hondrocita. Nakon 8 mjeseci za uzgoj u alginat gel zrelim kondrocitima ne gube metaboličku aktivnost i dalje sintetizirati tkivnospecifično kolagen tipa II i agrekan.
N. Tanaka i coauthors (1984) istraživali su svojstva difuzije različitih prirodnih molekula u alginatu i otkrili da molekule veće od 70 kD ne difuze kroz alginat. Dakle, kultivacija stanica u alginatu prikladna je za proučavanje regulacije biosinteze matrice i organizacije ECM. Dostupnost stanica obrađenih u alginatu omogućava ispitivanje učinka peptidnih regulatornih čimbenika i farmakoloških agensa na transkripcijske, posttranskripcijske i translacijske razine.
Hondrociti su također kultivirani u matrici kolagenskih vlakana I i II tipa. S. Nehrer i suradnici (1997), dok je operacija u pasa kondrocita proteoglikana osim kolagenom polimerni matriks sadrži kolagene različitih vrsta. Našli su značajne razlike u morfologiji biosintetskih funkcija kondrocitima uzgojenih u matriks kolagena sadrži kolagen tipa I i II. Stanice u matrici vrste kolagena II sohryanyali svoj sferični oblik, dok se kolagen tipa I imao fibroblasta poput morfologiju. Štoviše, u matrici tipa II kolagena, hondrociti su proizveli više glikozaminoglikana. J. Van Susante i sur. (1995) uspoređivali su svojstva hondrocita kultiviranih u alginat i gel kolagena (tip I). Autori su značajan porast broja stanica u kolagen gela, ali šesti dan uzgoja, stanice su izgubili karakterističan fenotip, pretvorio u fibroblasta nalik stanicama. U alginatnom gelu je zabilježeno smanjenje broja stanica, no hondrociti su zadržali svoj normalan fenotip. Količina kolagena gel proteoglikana po stanici bila je značajno veća nego u alginat, ali je uočeno smanjenje sinteze gel matriksa elementi počevši od 6. Dana uzgoja, dok se u sintezi alginat i dalje rasti.
Čvrsta trodimenzionalna matrica fibrina je prirodna supstanca koja podupire hondrocite koje se vagaju u njemu u diferenciranom fenotipu. 3D matrica fibrina također se može koristiti kao nosač za presađivanje kondrocita. Prednosti fibrina su odsutnost citotoksičnosti, sposobnost popunjavanja prostora, adhezivne sposobnosti. Histološkim i biokemijskim istraživanjima Autoren-diografii, elektronska mikroskopija pokazala je da su hondrociti u fibrin gel zadržati njihovu morfologiju, množe i stvaraju matricu i nakon 2 tjedna uzgoja. Međutim, G. Homminga i koautori (1993) su izvijestili da nakon 3 dana kulture počinje raspad fibrina, dedifferentirajući hondrociti napreduju.
Suspenzija kondrocita u umjetnom (sintetičkom) ECM-u
Implantati hrskavice za rekonstruktivnu ili ortopedsku kirurgiju mogu se dobiti uzgojem izoliranih hondrocita in vitro u sintetičkom biokompatibilnom matriksu.
Hondrociti kultiviranih poliglikolnih kiselina proliferiraju i održavaju normalnu morfologiju i fenotip unutar 8 tjedana. Kompleks kondrocita-poliglikolne kiseline sastoji se od stanica, glikozaminoglikana, kolagena i ima vanjsku kapsulu kolagena. Međutim, u takvim implantatima postoje dvije vrste molekula kolagena - I i II. Implantati dedifferentiated hondrocita niz prolaza imaju veću količinu glukozaminoglikana i kolagen nego u implantatima nediferenciranih primarnih hondrociti.
L. Freed et al (1 993b) u odnosu na ponašanje kondrocita kulturama ljudske i goveđe u vlaknastom poliglikolne kiseline (EQAP) i nesklad polilaktilovoy kiselini (PPLC). Nakon 6-8 tjedana uzgoja bikova hondrocita u HSVG ili PPLC, autori su promatrali staničnu proliferaciju i regeneraciju matriksa hrskavice. U HSBC-u su hondrociti bili sferni, smješteni u lacunima okruženi hrskavičnim matriksom. Nakon 8 tjedana uzgoja in vitro regenerirati tkivu sadržavati do 50% krutina (4% od mase stanica, 15% i 31% od glikozaminoglikana kolagena). U PPLK stanicama je oblik vretena, mala količina glikozaminoglikana i kolagena. U HSBC, rast stanica bio je 2 puta intenzivniji nego u PTCA. U uvjetima in vivo, hondrociti uzgojeni u HPVC i PPLC tijekom 1 do 6 mjeseci daju tkivo histološki sličnu hrskavici. Implantati sadržavali su glikozaminoglikane, tipa I i tip II kolagena.
Fetalni bikovi hondrociti uzgojeni su u poroznom hidrofobnom i hidrofilnom polietilenu visoke gustoće. Nakon 7 dana inkubacije u oba supstrata, stanice su zadržale sferični oblik, uglavnom koji sadržava kolagen tipa II. Nakon 21 dana uzgoja, pokazalo se da hidrofilna matrica sadrži više kolagena tipa II od hidrofobne matrice.
Tkivo hrskavice može se dobiti kultiviranjem u monosloju na Millicell-CM filtrima. Prethodno premazivanje filtera s kolagenom je neophodno za pričvršćivanje kondenzata. Histološki pregled kulture pokazuje nakupljanje kondrocita u ECG-u koji sadrži proteoglikane i tip II kolagena. Tip kolagena I u takvoj kulturi nije otkriven. Hondrociti u dobivenom hrskavom tkivu imaju sferični oblik, ali na površini tkiva oni su pomalo poravnati. Debljina novoformiranog tkiva povećao se s vremenom i ovisi o početnoj gustoći monosloja stanica. Pod optimalnim uvjetima kulture, debljina hrskavice dosegla je 110 μm, organizacija njezinih stanica i kolagena u površinskim i dubokim slojevima je slična onoj zglobne hrskavice. VKM sadrži oko 3 puta više kolagena i proteoglikana. Nakon 2 tjedna uzgoja zabilježeno je nakupljanje matriksa-sa, što je omogućilo izlučivanje tkiva iz filtera i koristi se za transplantaciju.
Sims i sur. (1996) proučavali su uzgoj hondrocita u polietilen oksid-gel kapsuliranoj polimernoj matrici koja omogućava prijenos velikog broja stanica injekcijom. Šest tjedana nakon ubrizgavanja u potkožno tkivo atimičnih miševa, formirana je nova hrskavica koja je morfološki karakterizirana bijelom opalescencijom sličnom hialinskoj hrskavici. Podaci iz histološke i biokemijske studije ukazuju na prisutnost aktivno proliferirajućih kondrocita, koji proizvode ECM.
Uzimanju
Ispitivanje hrskavičnog tkiva koristi se za proučavanje procesa ana- i katabolizma, homeostaze, resorpcije i popravka. Hondrociti u eksplozijama hrskavičnih tkiva podržavaju normalan fenotip i sastav ECM-a, sličan onima u zglobnoj hrskavici in vivo. Nakon 5 dana uzgoja u prisustvu seruma postiže se konstantna razina sinteze i prirodna razgradnja. Resorpcija može ubrzati kulturu tkiva, te u glavnoj kulturi uz dodatak seruma koristeći niz sredstvima, na primjer, IL-IB, TNF-a, bakterialnyhlipopolisaharidov, derivati retinoične kiseline ili aktivnim kisikovih radikala. Proučiti njegove oštećenja hrskavice popravak inducirana topivih medijatora upale (H 2 O 2, IL-1, TNF-a) ili fizičkog rupture matrice.
Metoda organotipskih kultura je model za proučavanje in vitro učinaka izoliranih vanjskih čimbenika na hondrocite i okolnu matricu. In vivo, hondrociti se rijetko nalaze u ECM-u i ne dolaze u kontakt. Kultura eksplanzijske zglobne hrskavice zadržava ovu strukturnu organizaciju, kao i posebnu interakciju između kondrocita i okolnog ekstracelularnog okruženja. Ovaj model se također koristi za proučavanje utjecaja mehaničkog stresa, farmakoloških tvari, čimbenika rasta, citokina, hormona na metabolizam hrskavice.
Druga prednost tkivne eksplancije tkiva je odsutnost štetnog gubitka kondrocita proteolitičkim enzimima ili mehaničkim čimbenikom koji je neizbježan kada se stanice izdvoje. Receptori i drugi membranski proteini i glikoproteini zaštićeni su od štetnih čimbenika.
Kultura kondrona
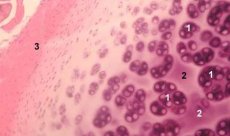
Hondron - strukturne, funkcionalna i metaboličke jedinica kod zglobne hrskavice koja se sastoji od kondrocita pericelularno matriksa i njegova kompaktan kapsule vlakna i odgovoran je za homeostazi matrice. Kondroni su mehanički izlučeni iz hrskavice i sakupljeni nekoliko sukcesivnih homogenizacijom male brzine. Izolirani iz zonama različite dubine hondrony hrskavice može se podijeliti u četiri kategorije: jedan hondron, twin hondrony, multiple (tri ili više) linearno postavljeni hondrony (kolona hondronov) hondronov zagušenja.
Jedan hondrony obično nalaze u sredini sloja netaknuta hrskavice, blizanca - na granici srednje i duboke slojeve linearno raspoređene multiple hondrony tipičan dubokih slojeva netaknutoj hrskavice. Konačno, klasteri kondrona sastoje se od nasumično organiziranih skupina pojedinačnih i uparenih kondija koji zadržavaju agregiranu državu nakon homogenizacije. Hondronov nakupine predstavljaju velike fragmente hrskavice, obično se sastoje od nekoliko radijalnih hondronov i kolagenskih vlakana, t, E. Tipična organizacija, koja je karakteristična za dubokih slojeva matrice. Kondroni su imobilizirani u prozirnoj agarozi, što omogućuje proučavanje njihove strukture, molekularnog sastava i metaboličke aktivnosti. Hondron sustav - agaroze smatra mikro model hrskavice, koji se razlikuje od tradicionalnog sustava kondrocita - agaroze koji čuva prirodna mikroklima, nema potrebe za obavljanje sinteze i montaža. Kultura kondrona je model za proučavanje interakcija stanica i matrice u zglobnoj hrskavici u normalnim i patološkim uvjetima.
 [22], [23], [24], [25], [26], [27]
[22], [23], [24], [25], [26], [27]
Kultura besmrtnih hondrocita
Za stvaranje trajnih staničnih linija koriste se rekombinantni DNA ili virusi koji sadrže onkogen koji mogu učiniti stanicu "besmrtnom". Besmrtni hondrociti imaju sposobnost beskrajne proliferacije, održavajući stabilan fenotip. F. Mallein-Gerin i suradnici (1995) pokazali su da je onkogen SV40T induciranu proliferaciju mišjih kondrocitima koji tako dalje stabilno eksprimiraju kolageni II, IX i XI vrste, kao i zajednički i agrekana vezanja proteina. Međutim, takva stanična linija stječe sposobnost sintetiziranja kolagena tipa I kada se kultiviraju u jednoslojnoj kulturi ili u agaroznom gelu.
W. Horton i koautori (1988) opisuju liniju besmrtnih stanica s niskom razinom ekspresije mRNA tipa kolagena II. Ove stanice su dobivene transformacijom s mišjim retrovirusom koji sadrži I-myc- i y-ra-onkoge. Ova vrsta stanica je jedinstveni model za proučavanje interakcija zglobne matrice u odsutnosti kolagena tipa II, kao i regulaciju sinteze tipa II kolagena.
Kultura kondropita s mutiranim ili izbrisanim genima prikladan je model za proučavanje njihove fiziološke funkcije. Ovaj model je posebno pogodan za proučavanje uloge određenih molekula u matriksa hrskavice organizacijama ili proučavanje učinaka različitih regulatornih čimbenika na metabolizam hrskavice. Hondrociti daljinski gena sintetizirati kolagen Tip IX kolagena fibrili šire nego što je normalno, što pokazuje da kolagen tipa IX regulira promjer fibrila. Kao što sam naveo u poglavlju 1, novoosnovanog kolagena genske mutacije vrsta kodiranja COLAI II u obitelji s primarnim generaliziranim osteoartritisa. Da bi se ispitao učinak mutanta kolagena tipa II na artikularnoj matrici R. Dharmrvaram et al (1997) izveden transfekcije ( „onečišćenja” stranih nukleinskih kiselina) defektnog COL 2 AI (arginin kod položaja 519 zamijenjen cisteinom) u humanim fetalnim kondrocitima in vitro.
Sustav kultura. U zglobu hrskavica djeluje zajedno sa stanicama drugih vrsta koje se nalaze u sinovijalnoj membrani, sinovijalnoj tekućini, ligamentima, subchondralnoj kosti. Na metabolizam hondrocita mogu utjecati različiti topljivi čimbenici sintetizirani ovim stanicama. Dakle, artritisna zglobna hrskavica uništava proteolitički enzimi i slobodni radikali koji se proizvode pomoću sinovijalnih stanica. Stoga su razvijeni modeli za proučavanje složenih interakcija između hrskavice i okolnih tkiva, koji se nazivaju kultura kulture.
S. Lacombe-Gleise et al (1995) su uzgojene zeca hondrocita i osteoblasta u sukulture sustavu (Costar), u kojem se stanice se rastave mikroporoznu membranu (0,4 um), omogućava razmjenu između dva tipa stanica bez izravnog kontakta. Ova studija je pokazala sposobnost osteoblasta da stimuliraju rast hondrocita putem topivih medijatora.
AM Malfait i koautori (1994) istražili su odnos između monocita periferne krvi i hondrocita. Taj model koristan za proučavanje procesa posredovanih upalnih citokina na artropatija (reumatoidni artritis, spondilitis seronegativne et al.). Autori modela odvojili su stanice proteinom vezujućim membranama s porama promjera 0,4 um. Istraživanje je pokazalo da lipopolisaharid-stimulirano monociti razrađen iFNO IL-1-a, koji inhibira sintezu hondrocita agrekana i doprinijelo razgradnje već sintetiziranih agrekanskih agregata.
K. Tada et al (1994) stvorio sukulture model u kojem endotelnih stanica u kolagen (I-tip), gel se stavi u unutarnjoj komori s vanjske komore od kojega ga kondrocitima smještena u filter s veličinom pora od 0,4 mikrona. U stanju potpune izolacije iz vanjske komore, ljudske endotelne stanice formirale su cijevi u kolagenskom gelu u prisutnosti EGF-a ili TGF-a. Uz simultano uzgoj obje vrste TGF stanica, ovisna tvorba cijevi pomoću endotelnih stanica je inhibirana. Hondrocita inhibicija ovog postupka djelomično je uklonjena anti-TGF-beta protutijelima. Može se pretpostaviti da TGF-beta proizvodi hondrociti koji smanjuju vaskularizaciju same hrskavice.
S. Groot i koautori (1994) istodobno su uzgojili hondrocite iz hipertrofnih i proliferativnih zona kosti 16 dana starog fetalnog miša s komadićima tkiva mozga. Nakon 4 dana kulture, promatrana je transdiferencijacija hondrocita u osteoblasta i nastanak osteoidnih oblika. Nakon 11 dana uzgoja, dio hrskavice zamijenjen je koštanim tkivom i koštana matrica je djelomično kalcificirana. Neki neuropeptidi i neurotransmiteri proizvedeni moždanim tkivom, utječu na metabolizam osteoblasta ili imaju receptore na njima. Među njima se može izolirati norepinefrin, vazoaktivni crijevni peptid, peptid povezan s galaktitima kalcitona, supstanca P i somatostatin. Uzgojeni s hondrocitima, dijelovi moždanog tkiva mogu proizvesti neke od tih čimbenika koji mogu inducirati proces transdiferenciranja hondrocita u osteoblaste.
[28], [29], [30], [31], [32], [33]
Utjecaj vanjskih faktora na kulturu kondrocita
Učinak napetosti kisika na metabolizam hondrocita
U većini slučajeva kulture hondrocita nastaju pod uvjetima atmosferske napetosti kisika. Ipak, dobro je poznato da in vivo hondrociti postoje pod hipoksičnim uvjetima, a napetost kisika varira različitim patološkim stanjima. Tijekom procesa sazrijevanja promatrane su značajne promjene u opskrbi krvi epifizama. Budući da se vaskularizacija razlikuje u različitim područjima ploče za rast, također se razlikuje i napetost kisika u njima. C. Brighton i R. Heppenstall (1971) pokazali su da je u pločici tibije kod zečeva napetost kisika u hipertrofnoj zoni manja nego u okolnoj hrskavici. Mjerenja nekih metaboličkih parametara pokazala su da hondrociti mogu brzo reagirati na lokalne promjene u koncentraciji kisika. Prije svega, uz nisku napetost kisika, njegova potrošnja kondrocita smanjuje. Uz smanjenje napetosti kisika od 21 do 0,04% povećava se iskorištavanje glukoze, povećava se aktivnost enzima glikolize i sinteza mliječne kiseline. Čak i uz malu napetost kisika, apsolutna količina ATP, ADP i AMP ostaje stabilna. Ovi podaci upućuju na usmjerenost metabolizma kondrocita radi maksimiziranja očuvanja energije. Ipak, sintetička aktivnost, a time i procesi obnove, mijenja se u uvjetima hipoksije.
Visoka napetost kisika također utječe na metabolizam hondrocita, uzrokujući smanjenje sinteze proteoglikana i DNA, degradaciju matriksa hrskavice. Ti učinci, u pravilu, popraćeni su proizvodnjom slobodnih radikala kisika.
Utjecaj koncentracije iona i osmotskog tlaka okoliša na funkciju hondrocita
U prirodnom koncentracije iona hrskavica se bitno razlikuje od ostalih tkiva: sadržaj natrija u ekstracelularni medij je 250 - 350 mmola, a osmolarnost - 350-450 mOsm. Za izolaciju hondrociti iz videorekordera i potom se inkubiraju u standardnom mediju (DMEM (Dulbecco minimalni esencijalni medij - Dulbecco minimalni esencijalni medij) osmolarnost - 250-280,7 mOsm) mijenja oštro okoline stanice. Osim toga, koncentracija kalcija i kalija u standardnim medijima je znatno niža nego kod domaćeg tkiva, a koncentracija aniona je znatno veća.
Dodavanje vodi saharoza povećati osmolarnost i inducira prolazno povećanje koncentracije intracelularnog H + i aniona kalcija u citosol. Takve unutarstanične promjene mogu utjecati na procese diferencijacije hondrocita i njihovu metaboličku aktivnost. J. Urban i suradnici (1993) su utvrdili da je uključivanje 35 8-sulfat i 3 H-prolina izoliranih kondrocitima inkubiraju u DMEM standardnom mediju za 2-4 sata, bila je samo 10% koje u izvornom tkivu. Intenzitet Sinteza je vrhunac kada je osmolarnost ekstracelularnog medija 350-400 mOsm u novo izoliranim hondrocita i eksplatanta hrskavice u. Osim toga, volumen kondrocita se povećao za 30-40% nakon stavljanja izoliranih stanica u standardni DMEM medij navedene osmolarnosti. Međutim, kada se uzgajaju hondrocite pod ne-fiziološkim osmolarnost za 12-16 sati, stanice se prilagodile novom okolišu smanjenjem intenziteta smicanja proporcionalna biosinteza osmolarnost izvanstanične medija.
P. Borgetti i suradnici (1995) istraživali učinak osmolarnost ekstracelularnog medija na rast, morfologija i biosintezu svinjski hondrociti. Autori su pokazali slične biokemijske i morfološke značajke hondrocita kultiviranih u mediju s osmolarnosti od 0,28 i 0,38 mosmola. Kada je uočeno 0,48 mOsm osmolarnost medija tijekom prvih 4-6 sati kulture smanjenje proliferacije stanica i sintezu proteina, ali nakon toga je došlo vraćanje tih parametara da bi na kraju stigli do kontrolne vrijednosti. Kada uzgajanje hondrocite u mediju s 0,58 mOsm osmolarnost stanice gube sposobnost za potporu fiziološkog intenziteta proliferacijske procese i nakon 6 tjedana značajno je smanjen broj hondrociti. Uz osmolarnost medija, 0,58 mosmola, opažena je duboka inhibicija sinteze proteina. Osim toga, kada se uzgajaju u mediju s osmolarnost mOsm 0,28-0,38 hondrociti zadržavaju fiziološko fenotip na višoj osmolarnost (mOsm 0,48-0,58) značajne promjene u morfologiji stanica, kao što je gubitak očituje karakterističan fenotip hondrociti konverziju u stanice slične fibroblastu, kao i gubitak stanica, sposobnost da se montiraju proteoglikani matriksa. Rezultati ove studije upućuju na sposobnost kondrocita da reagiraju na ograničene oscilacije osmolalnosti u ekstracelularnom okruženju.
Promjena koncentracije drugih iona također može utjecati na procese biosinteze u hondrocitima. Dakle, stupanj uključivanja 35 S (sulfata) povećava se polovicom s povećanjem koncentracije iona kalija od 5 mmol (koncentracija u standardnom DM DM mediju) do 10 mmol (koncentracija u VKM in vivo). Koncentracija kalcija ispod 0,5 mmola doprinijela je proizvodnji kolagena zrelim bikovim hondrocitima, dok je koncentracija od 1-2 mmol (koja odgovara koncentraciji u standardnom DM DM mediju) uzrokuje značajno smanjenje sinteze kolagena. Umjereno povećanje biosinteze zabilježeno je pri visokim razinama kalcija (2-10 mmol). Različiti kationi sudjeluju u vezanju hondrocita na VKM proteine. Dakle, magnezijev i manganski ioni osiguravaju vezanost s fibronektinom i kolagenom tipa II, dok kalcijevi ioni ne sudjeluju u vezivanju kondrocita u proteine. Prema tome, rezultati opisanih istraživanja ukazuju na utjecaj promjene izvanstaničnih iona kalijuma, natrija, kalcija i osmolarnosti medija na biosintetsku funkciju hondrocita inkubiranih u standardnim medijima.
Utjecaj mehaničkog stresa na metabolizam hondrocita
Imobilizacija zglobova uzrokuje reverzibilnu atrofiju hrskavice, što ukazuje na potrebu mehaničkih podražaja za normalni tijek metaboličkih procesa u ECM-u. U većini slučajeva, korišteni modeli stanične kulture postoje pod normalnim uvjetima atmosferskog tlaka. M. Wright i koautori (1996.) pokazali su da mehanička okolina utječe na metabolizam hondrocita, odgovor stanica ovisi o intenzitetu i učestalosti kompresijskog opterećenja. Eksperimenti s opterećenjem na netaknutim dijelova biljaka iz zglobne hrskavice in vitro pokazala su smanjenje sinteze proteina i proteoglikana pod statičkim opterećenjem, dinamičko opterećenje, dok poticanje tih procesa. Točni mehanizmi za provedbu mehanička opterećenja na učinke hrskavice kompleksa, a vjerojatno povezane s naprezanje stanice na hidrostatički tlak, osmotski tlak, električnog potencijala i receptore na staničnoj površini za matrične molekula. Za proučavanje učinka svakog od ovih parametara potrebno je stvoriti sustav u kojem se jedan parametar može samostalno mijenjati. Na primjer, kultura eksplantacije nije pogodna za proučavanje stanične deformacije, ali se može koristiti za proučavanje ukupnog učinka pritiska na metaboličku aktivnost kondrocita. Kompresija hrskavice dovodi do deformacije stanica, a također uz pojavu hidrostatskog tlaka, gradijenta električnog potencijala, i protok tekućine promjene fizikalno faktorima kao što je sadržaj vode u matrici, gustoći električnog naboja, razine osmotskog tlaka. Stanična deformacija može se proučavati pomoću izoliranih hondrocita uronjenih u agarozu ili kolagenski gel.
Razvijeno je nekoliko sustava za proučavanje utjecaja mehaničke stimulacije na kulturu kondrocita. Neki istraživači koriste takve sustave u kojima se tlak primjenjuje na kulture stanica kroz plinovitu fazu. Na primjer, JP Veldhuijzen et al (1979), koristeći tlak iznad atmosferskog 13 kPa, na niskoj frekvenciji (0.3 Hz), tijekom 15 minuta, uočeno povećanje sintezu cAMP i proteoglikana i spuštanje sintezu DNA. R. Smith et al (1996) pokazali su da je intermitentna izlaganje primarnim kulturama hondrociti bik hidrostatskog tlaka (10 MPa), na 1 Hz tijekom 4 sata uzrokovala porast sinteze agrekana i kolagen tipa II, a konstantni tlak nije imao učinka na ove procese. Korištenje sličnog sustava M. Wright i sur (1996) su izvijestili da je ciklička pritisak na staničnoj kulturi je povezana s hiperpolarizacije u staničnoj membrani hondrocite i aktiviranjem Ca 2+ ovisnog kalij kanala. Dakle, učinci cikličkog tlaka su posredovani ionskim kanalima, aktiviranim istezanjem, u membranskoj kondukciji. Odziv hondrocita prema hidrostatskom tlaku ovisi o uvjetima stanične kulture i učestalosti primijenjenog opterećenja. Tako, ciklički hidrostatski tlak (5 MPa) smanjuje ugradnju sulfata u monosloju kondrocita na frekvenciji od 0,05, 0,25 i 0,5 Hz, a za frekvencije iznad 0.5 Hz inkluzijskog sulfata eksplatanta hrskavice povećava.
Bushmann M. I suradnici (1992) su izvijestili da hondrociti u agaroznom gelu Mr. Mijenjati biosintezu odgovor na statička i dinamička mehanička opterećenja, kao i kultivirani netaknuti organ. Autori su otkrili da mehaničko opterećenje stvara hiperosmotski poticaj nakon čega slijedi smanjenje pH u hondrocitima.
Učinak mehaničkog istezanja može se proučavati na kulturi stanica uronjenih u gel. Sila istezanja može se stvoriti pomoću računala kontroliranog vakuuma. Kada je sustav u određenom stupnju vakuumu dno Petrijevoj zdjelici sa kulturom stanica proširi poznata količina, maksimalne deformacije na rubovima dna čaške minimuma u središtu. Istezanje se prenosi i uzgaja u Petrijevom jelu kondrocita. Tom metodom, Holm-Vall K. Et al (1995) su pokazali da su uzgojene u kolagen (II) tipa gel hondrosarkom stanica povećava ekspresiju mRNA i 2 -integrina. 2 p r integrina može vezati na kolagenom tipa II. Smatra se mehanoreceptorom, jer interakcionira s proteinima koji vežu aktin, povezujući time ECM i citoskelet.
Učinak pH na metabolizam kondrocita
PH intersticijalne tekućine ECM hrskavog tkiva je kiselljije nego u drugim tkivima. A. Maroudas (1980) određuje pH zglobne hrskavice na 6.9. W. Diamant i koautori (1966) pronašli su pH vrijednosti od 5,5 u patološkim uvjetima. Poznato je da hondrociti žive na niskom PO2, što ukazuje na važnu ulogu glikolize (95% ukupnog metabolizma glukoze) u metabolizmu ovih stanica; glikoliza je praćena proizvodnjom velike količine mliječne kiseline.
Uz acidifikaciju okoliša kod proizvoda glikolize, same su komponente matrice vrlo važne. Veliki broj fiksne negativnog naboja na ekstracelularne proteoglikana modificira ionski sastav: postoji visoka koncentracija slobodnih kationa (npr H +, Na +, K + ) i niske koncentracije aniona (npr O2, NPHS). Osim toga, pod utjecajem mehaničkog opterećenja voda se protjeruje iz ECM-a, što dovodi do povećanja koncentracije fiksnih negativnih naboja i privlačenja više kationa u matricu. To je popraćeno smanjenjem pH izvanstaničnog medija koji utječe na intracelularni pH, čime se modificira metabolizam hondrocita. R. Wilkin i A. Hall (1995) proučavali su učinak pH izvanstaničnog i unutarstaničnog medija na biosintezu matrice pomoću izoliranih bikondrocita. Primijetili su dvostruku modifikaciju sinteze matrice s padom pH. Maleno povećanje pH vrijednosti (7.4
[34], [35], [36], [37], [38], [39], [40]
Učinak pripravka medija za uzgoj na metabolizam hondrocita
Medij za uzgoj hondrocita mora odgovarati eksperimentalnim uvjetima. Posljednjih godina, teleći serum je korišten za optimizaciju uvjeta kulture. Međutim, kada se koristi serum, treba uzeti u obzir nekoliko važnih točaka:
- vanjski rast stanica s periferije tkiva u kulturama organa,
- varijabilnost sastava sera različitih serija,
- prisutnost nepoznatih komponenti u njima,
- povećan rizik od smetnji, artefakti u proučavanju utjecaja različitih bioloških čimbenika na metaboličku aktivnost stanica.
Primjer ovog potonjeg je proučavanje učinka EGF-a na hrskavaste hondrocite kod štakora. EGF stimulira ugradnju 3 timidinom i povećati sadržaj DNA u kulturi. Taj je učinak bio izraženiji pri niskim koncentracijama seruma (<1%), ali pri visokim koncentracijama (> 7,5%) efekt je nestao.
Dobro je poznato da su razine sinteze i degradacije u DMEM obogaćene telećim serumom značajno povećane u odnosu na in vivo uvjete. Razlike između metabolizma in vivo i in vitro mogu biti uzrokovane razlikama između sinovijalne tekućine i okoliša u kojem se uzgajaju stanice. D. Lee i sur (1997) su kultivirane hondrociti junaca agaroznom pomoću hranjivi medij sadrži DMEM, obogaćen s 20% telećeg seruma i veliki broj alogene normalnog sinovijalnoj tekućini. Prisutnost sinovijalne tekućine u mediju potaknula je povećanje broja proteoglikana, do 80% ukupne količine sinovijalne tekućine. Dobiveni rezultati ukazuju da sinovijalna tekućina u kulturi inducira brzinu metabolizma sličnu onima in vivo, s visokom razinom sinteze glikozaminoglikana i niskom razinom stanične diobe.
G. Verbruggen i suradnici (1995) pokazali su da je sinteza 35 S-arrpeKaHa ljudske hondrocite uzgojene u DMEM agarozom u bez seruma je 20-30% razine promatrane sinteze u DMEM, uz dodatak 10% telećeg seruma. Autori su utvrdili u kojoj mjeri IGF-1, IGF-2, TGF-P ili inzulin vraćaju proizvodnju agrekana u mediju bez seruma. Autori su zaključili da su 100 ng / ml inzulina, IGF-1 i IGF-2 djelomično smanjene sinteze agrekana na 39-53% od kontrolnih nivoa. S kombinacijom tih čimbenika nije utvrđeno sinergističko ili kumulativno pojavljivanje. Istovremeno, 10 ng / ml TGF-P u prisustvu 100 ng / ml inzulina stimulira sintezu agrekan na 90% ili više od referentne razine. Konačno, serum transferrin, sam ili u kombinaciji s inzulinom, nije utjecao na sintezu agrekana. Kada je teleći serum zamijenjen goveđim serumskim albuminom, agregatni sadržaj agregata značajno je smanjen. Obogaćivanje medija za inzulinsku kulturu, IGF ili TGF-P djelomično je obnovila sposobnost stanica da proizvode agrekanske agregate. U tom slučaju, IGF-1 i inzulin su u stanju održavati homeostazu u staničnim kulturama. Nakon 40 dana u kulturi mediju obogaćenom s 10-20 ng / ml IGF-1, proteoglikana Sinteza se održava na istoj razini ili čak i više u odnosu na medij koji sadrži 20% telećeg seruma. Katabolički procesi polako kreće u mediju obogaćenom sa IGF-1 u odnosu na mediju obogaćenom s 0,1% otopinom albumina, ali nešto brže u mediju, uz dodatak 20% seruma. U dugotrajnim kulturama, 20 ng / ml IGF-1 održava stabilno stanje stanica.
D. Lee i sur (1993), dok učinak pripravka medija kulture (DMEM, DMEM + 20% telećeg seruma, DMEM + 20 ng / ml IGF-1) na sintezu DNA u kulturi eksplanata hrskavice, jednoslojne kulture u suspenziji u agaroza , Kada su uzgojeni agarozom u prisustvu seruma, autori su promatrali tendenciju skupljanja hondrocita u velike skupine. Stanice su uzgojene bez seruma i IGF-1, zadržavaju kružni oblik u agaroze, prikupljeni su u malim grupama, ali ne čine velike nakupine. U monosloju, sinteza DNA bila je znatno veća u mediju koji sadrži serum nego u mediju obogaćenim s IGF-1; Sinteza DNA u potonjem bila je znatno veća nego u neograđenom okolišu. Kada kultiviranje hondrocite u suspenziji u agaroza unconcentrated mediju i u mediju sa IGF-1, bez razlike u sintezi DNA. Istovremeno mulj kultiviranje hondrocite u agarozom u mediju nadopunjenom sa serumom, praćen je povećanjem ugradnjom radionukleotid 3 timidinom usporedbi s drugim okruženjima.
Vitamin C je potreban za aktivaciju enzima uključenih u formiranje stabilne spiralne strukture kolagenskih fibrila. Hondrociti, deficijentni s obzirom na askorbinsku kiselinu, sintetiziraju pod-hidroksilirane nehelične prekursore kolagena koji se polagano izlučuju. Uvođenje askorbinske kiseline (50 ug / ml) uzrokuje hidroksiliranje tipa II i IX kolagena i njihovo lučenje u normalnim količinama. Dodavanje vitamina C nije utjecalo na razinu sinteze proteoglikana. Posljedično, izlučivanje kolagena regulirano je neovisno od izlučivanja proteoglikana.