
Virus humane imunodeficijencije (HIV)
Posljednji pregledao: 23.04.2024

Svi iLive sadržaji medicinski se pregledavaju ili provjeravaju kako bi se osigurala što je moguće točnija činjenica.
Imamo stroge smjernice za pronalaženje izvora i samo povezujemo s uglednim medijskim stranicama, akademskim istraživačkim institucijama i, kad god je to moguće, medicinski pregledanim studijama. Imajte na umu da su brojevi u zagradama ([1], [2], itd.) Poveznice koje se mogu kliknuti na ove studije.
Ako smatrate da je bilo koji od naših sadržaja netočan, zastario ili na neki drugi način upitan, odaberite ga i pritisnite Ctrl + Enter.
Sindrom stečene imunodeficijencije izoliran je kao posebna bolest 1981. Godine u Sjedinjenim Državama, kada je u mnogim mladim ljudima ozbiljne bolesti uzrokovane mikroorganizmima koji nisu patogeni ili neznatno patogeni za zdrave ljude. Istraživanje imunološkog statusa pacijenata otkrilo je značajno smanjenje broja limfocita općenito i osobito T-pomoćnika. Ovo se stanje naziva AIDS (sindrom stečene imunodeficijencije u Engleskom - Sindrom stečene imunodeficijencije ili AIDS). Metoda infekcije (seksualni kontakt, kroz krv i njegove pripravke) ukazuje na zaraznu prirodu bolesti.
Uzrok uzročnika AIDS-a 1983. Godine je samostalno otkrio francuski L. Montagnier, koji ga je nazvao LAV Lymphoadenopathy Associated Virus), dok je otkrivao pacijenta s limfadenopatijom; i američki R. Gallo, koji je nazvao virus HTLV-III (engleski humani T-limfotropni virus III): prethodno je pronađen limfotropni virusi I i II.
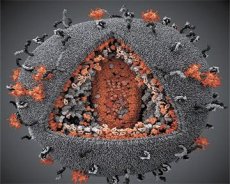
Usporedba svojstava virusa LAV i HTLV-III pokazala je njihov identitet, pa je 1986. Virus nazvan HIV (virus ljudske imunodeficijencije ili HIV) kako bi se izbjegla zbrka. HIV je sferni, promjer je 110 nm. Omotnica virusa ima oblik poliedara koji se sastoji od 12 pentagona i 20 heksagona. Molekula glikoziliranog proteina gpl20 nalazi se u središtu i uglovima svakog šesterokuta (broj 120 označava molekularnu težinu proteina u kilodaltonima). Ukupno je 72 molekule gpl20 na površini viriona, od kojih je svaka povezana s intramembranskim proteinom gp41. Ti proteini zajedno s dvostrukim lipidnim slojem tvore superkapsušu (membranu) viriona.
Proteini gpl20 i gp41 nastaju kao posljedica rezanja stanične proteaze prekursorskog proteina Env. Protein gp41 tvori "klin" kralježnice, koji je povezan s citoplazmatskom domenom s matričnim proteinom p17MA odmah ispod membrane. Molekule p17, koje djeluju u interakciji sa sazreljem viriona, formiraju icosaedar koji leži ispod ljuske.
U središnjem dijelu viriona, protein p24 tvori konusni kapsid. Suženi dio kapsida uz sudjelovanje proteina pb povezan je s omotom viriona. Unutar kapsida postoje dvije identične molekule viralne genomske RNA. Oni su vezani na svojim 5 'krajevima na nukleozapidni protein p7NC. Ovaj protein je zanimljivo da ima dva aminokiselinska ostatka (motiv), cistein-bogatih i histidin atom i koji sadrži Zn - oni se nazivaju „cink 'finger”, kao što su hvatanje molekule genomske RNA za uključivanje u viriona formirana. Kapsid također sadrži tri enzima. Reverzne transkriptaze (RT) ili pol-kompleks obuhvaća reverznu transkriptazu, RNaze H i DNA-ovisne DNA polimeraze. Revertaza je prisutna kao p66 / p51 heterodimer. Proteaza (PR) - pI, započinje i ostvaruje proces sazrijevanja viriona. Integracija (IN) - p31, ili endonukleaza, osigurava uključivanje proviralne DNA u genom stanice domaćina. Kapsid također sadrži molekulu sjemena RNA (tRNAl "3).
RNA gena u stanici je prevedena reverznom transkriptazom u DNA genomu (DNA-provirus) koji se sastoji od 9283 nukleotidnih parova. Ograničeno je na lijevu i desnu stranu takozvanih dugotrajnih ponavljanja, ili LTR (engl. Long terminal repeat): S'-LTR - lijevo i Z'-LTR - s desne strane. LTR sadrži 638 parova nukleotida.
HIV genom se sastoji od 9 gena, od kojih su neki preklapani (ima nekoliko okvira za čitanje) i ima eksoninsku strukturu. Oni kontroliraju sintezu 9 strukturnih i 6 regulacijskih proteina.
LTR vrijednost za virusni genom je da sadrže sljedeće regulacijske elemente koji kontroliraju njegovo funkcioniranje:
- transkripcijski signal (promotorsko područje);
- signal za dodavanje poli-A;
- Signal hvatanja;
- integracija signala;
- pozitivni regulatorni signal (TAR za TAT protein);
- element negativne regulacije (NRE za NEF protein);
- mjesto vezanja sjemene RNA (tRNA ™ 3) za sintezu minus lanca DNA na kraju 3 '; signal na 5'-kraju LTR, koji služi kao primer za sintezu plasnog lanca DNA.
Osim toga, LTR sadrži elemente uključene u regulaciju spajanja mRNA, pakiranje molekula vRNA u kapsid (Psi element). Konačno, kada transkribiraju genom u dugim mRNA-ima, dva su signala generirana za REV protein, koji prebacuju sintezu proteina: CAR za regulacijske proteine i CRS za strukturne proteine. Ako se REV protein veže na CAR, sintetizirani su strukturni proteini; ako je odsutan, sintetizirani su samo regulatorni proteini.
U regulaciji genoma virusa, slijedeći regulacijski geni i njihovi proteini igraju posebno važnu ulogu:
- TAT protein koji provodi pozitivnu kontrolu reprodukcije virusa i djeluje kroz regulatorni TAR mjesto;
- proteina NEV i VPU, provodeći negativnu kontrolu reprodukcije kroz NRE mjesto;
- protein REV, provodeći pozitivnu negativnu kontrolu. REV protein kontrolira rad gena gage, pol, env i provodi negativnu regulaciju spajanja.
Tako je reprodukcija HIV-a pod trostrukom kontrolom - pozitivna, negativna i pozitivno-negativna.
VIF protein određuje infektivnost novosintetiziranog virusa. Ona je vezana za kapsidni protein p24 i prisutan je u virionu u količini od 60 molekula. NEF protein je prikazan u virionu malim brojem molekula (5-10), eventualno povezanim s omotnicom.
VPR protein inhibira stanični ciklus u fazi G2, u koji su uključeni u transport preintegratsionnyh kompleksa u jezgru stanice i aktivira određene virusne i stanične gene, povećava učinkovitost replikacije virusa u monocitima i makrofagima. Mjesto proteina VPR, TAT, REV, VPU u virionu nije utvrđeno.
Pored vlastitih proteina, sastav virionske membrane može uključivati neke proteine stanice domaćina. Proteini VPU i VPR su uključeni u regulaciju reprodukcije virusa.
Antigene varijante virusa humane imunodeficijencije (HIV)
Virus humane imunodeficijencije (HIV) vrlo je varijabilan. Čak i iz organizma jednog bolesnika mogu se izolirati sojevi virusa koji se značajno razlikuju u antigenskim svojstvima. Takvu varijabilnost potiče intenzivno uništavanje CD4 + stanica i moćan odgovor antitijela na HIV infekciju. Pacijenti iz Zapadne Afrike imaju novi oblik HIV-a, biološki bliski HIV-1, ali imunološki različit od njega, HIV-2. Homologija primarne strukture genoma ovih virusa je 42%. DNA-provirus HIV-2 sadrži 9671 bp, a njegov LTR-854 bp. HIV-2 je kasnije izoliran u drugim regijama svijeta. Nema križnog imuniteta između HIV-1 i HIV-2. Dva su velika oblika HIV-1 poznata: O (Outlier) i M (Major), potonji su podijeljeni u 10 podtipova (AJ). U Rusiji cirkuliraju 8 podtipova (AH).
 [
[Mehanizam interakcije HIV-a sa stanicom
Nakon što je prodrla u tijelo, virus prvi napada stanice koje sadrže specifični CD4 receptor. Ovaj receptor ima veliki broj T-pomoćnika, u manje makrofagima i monocitima, posebno virusima osjetljivim T-pomoćnicima.
Virus humane imunodeficijencije (HIV) prepoznaje CD4 receptore svojim gpl20 proteinom. Proces interakcije HIV-a sa stanicom prolazi prema sljedećoj shemi: adsorpcija posredovana receptorom -> granična fovea -> granična vezikula -> lizosom. U njemu se virionska membrana spaja s lizosomskom membranom, a nukleokapsid, oslobođen iz superkapsida, ulazi u citoplazmu; na putu do jezgre je uništena, a genomska RNA i povezane komponente jezgre se otpuštaju. Tada reverzna transkriptaza sintetizira minus lanac DNA na virion RNA, tada RNA-ase H uništava virion RNA, a virusna DNA polimeraza sintetizira plus-lanac DNA. Na krajevima DNA-provirusa nastaju 5'-LTR i 3'-LTR. DNA-provirus može biti u jezgri neko vrijeme u neaktivnom obliku, ali prije ili kasnije integrira se uz pomoć svoje integraze u kromosom ciljne stanice. U njemu, provirus je u neaktivnom stanju dok ovaj T-limfocit ne aktivira mikrobni antigeni ili druge imunokompetentne stanice. Aktivacija transkripcije stanične DNA regulirana je posebnim nuklearnim faktorom (NF-kB). To je protein koji veže DNK i proizvodi se u velikim količinama tijekom aktivacije i proliferacije T-limfocita i monocita. Taj se protein veže na specifične sekvence stanične DNA i slične sekvence LTR DNA-provirusa i potiče transkripciju i stanične DNA i DNA-provirusa. Pokretanjem transkripcije DNA-provirusa, on prelazi virus iz neaktivnog stanja u aktivnu, a time i uporni zarazu, u produktivnu. Ostanite provirus u stanju neaktivnosti može trajati jako dugo. Aktivacija virusa kritički je trenutak u interakciji sa stanicom.
Od trenutka kada virus uđe u stanicu počinje razdoblje infekcije HIV-om, virus koji može trajati 10 godina ili više; i od aktivacije virusa počinje bolest - AIDS. Uz pomoć svojih regulatornih gena i njihovih proizvoda, virus počinje aktivno razmnožavati. TAT-protein može povećati brzinu reprodukcije virusa 1000 puta. Transkripcija virusa je složena. To uključuje formiranje i pune i duljine mRNA, spliciranje mRNA i daljnju sintezu strukturnih i regulatornih proteina.
Sinteza strukturnih proteina dolazi kako slijedi. Početku sintetizira poliprotein prekursora Pr55Gag (protein sa m. M. 55 kD). On sadrži četiri glavna područja: matriks (MA), ovojnice (CA), nukleokapsid (NC) i PE domena, od kojih je rezultat smanjenje Pr55Gag virusnih proteaza (to samovyrezaetsya iz drugog proteina prekursora - gag-pol) nastaju pojedinačno strukturnih proteina P17 , p24, p7 i pb. Obrazovanje Pr55Gag poliprotein - glavni uvjet za formiranje virusnih čestica. To je taj protein određuje programa virusne morfogeneze. On uključuje stupnjeve uzastopno gag poliprotein transport u staničnom membranom i njegove interakcije s protein-protein interakcije na formiranje čestica virusa i njegova pupanje. Pr55Gag je sintetiziran na slobodnim polibosomima; Proteinske molekule transportiraju na membranu, koja je usidrena njihovi hidrofobnim područjima. Glavna uloga u stvaranju nativnu konformaciju proteina ima gag-CA domenu. Prekidač NC domena daje (pomoću „cinkovih prstiju”) 2 molekule genomske RNA u virusnu česticu ploču. Poliprotein prva molekula dimerizira interakcijom matriks domena. Zatim dimeri Spojeni u heksamernih (6 jedinica) kompleksa interakcijom domene CA i NC. Konačno, heksamera, povezuju bočne površine čine nezrelih virione sferični oblik, u kojem sadrže viralne genomske RNA snimljene NC-domene.
Drugi protein prekursor Prl60Gag-Pol (protein sa m. M. 160 kDa), sintetiziran kao rezultat pomicanja čitanja okvira ribosoma za prevođenje Z'-kraju gag gena u područje neposredno prije područje kodiranja RB proteina. Ovaj Gag-Pol poliprotein sadrži nepotpunu sekvencu Gag proteina (1 - 423 aminokiseline) i Pol sekvence, koje uključuju PR, RT i IN domene. Molekule poliproteina Gag-Pol se također sintetiziraju na slobodnim polibosomima i prenose se u plazmatsku membranu. Poliprotein Prl60Gagpol sadrži sve lokacije intermolekularnih interakcija koje su svojstvene poliprotein Gag i membranskim veznim mjestima. Stoga, molekule iz poliproteinskog gag-pol osigurača s membranom i, zajedno s gag molekule uključuju virione tvore, što bi moglo rezultirati aktivna proteaze i proces sazrijevanja virion počinje. HIV-1 Proteaza visoko aktivne samo kao dimer, i nje od samovyrezaniya Prl60Gag-Pol zahtjeva dimerizaciju ovih molekula. Sazrijevanje viriona je da otpuštena aktivna proteaza smanjuje prl60Gag-Pol i Gag55 u prepoznatljiva mjesta; nastaju proteini p17, p24, p7, p6, revertaza, integraza i njihovo povezivanje u viralnu strukturu.
Env protein sintetiziraju na ribosomima povezane s membranama endoplazmatski retikulum, glikozilirani, a zatim se reže u staničnu proteaze gp120 i gp41 i transportiraju na površinu stanice. U tom slučaju, gp41 prožima membranu i veže se na matrične domene Gag proteinske molekule povezane s unutarnjom površinom membrane. Taj odnos postoji u zrelom virionu.
Tako, montaža virusnih čestica je agregacija prekursora proteina i RNA molekula povezanih na plazma membranu stanice domaćina, formiranje nezrelih viriona i njihovo oslobađanje pupanjem iz površine stanice. Kada se pupi, virion se okružuje staničnom membranom, u kojoj su ugrađene molekule gp41 i gp120. Tijekom pupanja ili moguće nakon otpuštanja viriona sazrijevanja nastaje, koja se provodi uporabom virusnih proteaza se proteolitički rezanje Pr55Gag prekursora proteina i Prl60Gag-Pol virus zrele proteina i njihova povezanost s određenim strukturnim kompleksa. Vodeću ulogu u morfogeneze virusa ima poliproteinski prekursor Pr55Gag koji organizira i montažu nezrelih virion; Proces sazrijevanja popunjava specifična virusna proteaza.
Uzroci imunodeficijencije
Jedan od glavnih uzroka imunodeficijencije zaraze HIV-om je masivan gubitak T-helper stanice. Pojavljuje se zbog sljedećih događaja. Prvo, da je zaražena virusom T-pomoćničkih stanica umire zbog apoptoze. Smatra se da pacijenti s virusom AIDS replikacije, apoptoze i smanjenje broja T-pomoćnih stanica međusobno. Drugo, T-stanice ubojice prepoznaju i uništavaju T-stanica inficiranih virusom ili nosi adsorbira gpl20 molekule, kao inficiranim s virusom i virusom inficiranih T-pomoćnih stanica, koje tvore symplasts (sincicij) koja se sastoji od nekoliko desetaka stanica (kao dio oni umiru kao rezultat umnožavanja virusa u njima). Zbog uništenja velikog broja T-helper stanice se javlja ekspresija smanjenje membrana receptora u B-limfocita s IL-2, poremećena sinteza raznih interleukina (faktore rasta i diferencijacije B-limfocita -, IL-4, IL-5, IL-6, i drugi), zbog čega se krši funkcija sustava T-ubojica. Pojavljuje se suzbijanje aktivnosti komplementarnih i makrofaga. Virus-om makrofagi i monociti umrijeti, ali oni nisu u mogućnosti ukloniti virus iz tijela. Konačno, s obzirom na strukturne sličnosti i antigenske s receptorima gpl20 neke epitelne stanice organizma (uključujući trofoblaste receptorskog posredovanju prijenos HIV transplantata) sintetiziran antiretseptornyh protutijela sa širokim spektrom djelovanja. Takva antitijela mogu blokirati različite stanične receptore i komplicirati tijek autoimunih poremećaja bolesti. Posljedica infekcije HIV-om je porazio sve glavne dijelove imunološkog sustava. Takvi bolesnici postaju bespomoćni protiv širokog spektra mikroorganizama. To dovodi do razvoja oportunističkih infekcija i neoplastičnih bolesti. Za zaraženih HIV-om imaju povećan rizik od razvoja raka s najmanje tri vrste Kaposijev sarkom; karcinomi (uključujući rak kože); B-stanični limfom, koji proizlazi iz maligne transformacije B-limfocitima. Međutim, HIV ima samo limfocit, ali i neurotropski. Prodire u stanice CNS (astrociti) bilo endocitoze izazvane pomoću receptora i fagocitoza astrociti s virusom inficiranim lymphoblasts. U interakciji virusa s astrociti su formirane symplasts pogoduju širenju patogena međustanične kanala. U makrofaga i monocita, virus može trajati duže vrijeme, tako da oni služe kao rezervoar i njegovih distributera u tijelu, biti u stanju prodrijeti u svim tkivima. Zaražene makrofagi igra glavnu ulogu u rasipanjem HIV u središnjem živčanom sustavu i poraza. U 10% bolesnika s primarnim kliničkih sindroma povezanih s CNS-a i prikazanih u obliku demencije (demencija). Tako, za oboljele od HIV infekcije je karakterizirana 3 skupine bolesti - oportunističkih infekcija, tumorskih oboljenja i CNS.
Epidemiologija HIV infekcije
Izvor zaraze HIV-om je samo osoba - bolesna ili nosač virusa. Virus humane imunodeficijencije (HIV) se nalazi u krvi, sjemenu i cervikalnoj tekućini; u majkama - u majčinom mlijeku. Infekcija se događa seksualno, kroz krv i njene lijekove, kao i od majke do djeteta prije isporuke, tijekom i poslije porođaja. Slučajevi infekcije virusom kroz hranu, piće i kroz ubode insekata nisu poznati.
Ovisnost o drogi pridonosi širenju AIDS-a. HIV infekcija se povećava svake godine. Prema WHO-u, od 1980. Do 2000. Godine, 58 milijuna ljudi zaraženo je HIV-om. Samo u 2000, 5,3 milijuna ljudi zaraženo je u svijetu, a 3 milijuna ljudi je umrlo od AIDS-a. U Rusiji je od 1. Siječnja 2004. Registrirano 264.000 HIV pozitivnih osoba. Polovica zaraženih HIV-om umre u roku od 11-12 godina od infekcije. Početkom 2004. Godine, od svakih 100.000 ruskih građana, oko 180 je živjelo s dijagnozom "infekcije HIV-om". Predviđeno je da će na ovoj razini učestalosti ukupnog broja osoba zaraženih HIV-om u Rusiji do 2012. Godine biti 2,5-3 milijuna ljudi. Složenost borbe protiv HIV infekcije ovisi o nizu razloga: prvo, ne postoje učinkovite metode njegovog liječenja i specifične prevencije; Drugo, razdoblje inkubacije za HIV infekciju može premašiti 10 godina. Njeno trajanje ovisi o trenutku aktivacije T-limfocita i DNA-provirusa koji se nalazi u njegovom kromosomu. Još nije jasno je li svaki virus zaražen AIDS-om osuđen ili je vjerojatno da će imati dugotrajni virus bez bolesti (što izgleda malo vjerojatno). Konačno, postoji nekoliko ljudskih virusa imunodeficijencije (HIV-1, HIV-2), antigenskih razlika između kojih se sprječava stvaranje križnog imuniteta. Detekcija virusa imunodeficijencije majmuna (SIV) osvjetljava porijeklo HIV-a. SIO za organiziranje genoma sličan je HIV-u, ali se značajno razlikuje u nukleotidnoj sekvenci. HIV-2 serološki zauzima srednji položaj između HIV-1 i SIV, a nukleotidni slijed bio je bliži SIV-u. U tom pogledu, VM Zhdanov je sugerirao da virusi HIV-1, HIV-2 i SIV potječu od zajedničkog pretka. Moguće je, prema R. Gallo, da je jedan od SIV-ova nekako ušao u ljudsko tijelo, gdje je podvrgnut nizu mutacija, što je rezultiralo HIV-1, HIV-2 i drugim oblicima.
Simptomi infekcije HIV-om
Virus humane imunodeficijencije karakterizira određena svojstva na kojima patogeneza bolesti u velikoj mjeri ovisi. Virus ima vrlo visok stupanj reprodukcije, određen njegovim regulatornim elementima (5,000 viriona se sintetiziraju unutar 5 minuta u aktivnoj fazi). Zbog prisustva fuzijskog proteina (gp41), virus potiče stvaranje opsežnih sincicijskih struktura uslijed fuzije inficiranih i neinficiranih T-pomoćnika, što rezultira njihovom masovnom smrću. Molekule velikih molekula gpl20 slobodno cirkuliraju u krvi i vežu se na receptore neinfektiranih T-pomoćnika, kao rezultat toga što ih T-ubojice prepoznaju i uništavaju. Virus se može širiti kroz međustanične kanale od stanice do stanice, u tom slučaju postaje manje dostupan protutijelima.
Klinički kriteriji za HIV infekciju
HIV odraslih uspostaviti ako imaju najmanje dvije ozbiljne simptome u kombinaciji s barem jednim simptom maloljetnika u nedostatku drugih poznatih uzroka imunodeficijencije (rak, kongenitalne imunodeficijencije, teške gladi, i tako dalje. P.). Ozbiljni simptomi uključuju:
- gubitak težine za 10% ili više;
- produljena vrućica, povremena ili uporni;
- kronični proljev.
Manje simptomi uključuju uporan kašalj, dermatitis, generalizirani ponavljajućeg herpes zoster, kandidijaza usne šupljine i ždrijela, kronična herpes simplex, generaliziranom limfadenopatijom. Dijagnoza AIDS-a se provodi uz prisutnost samo Kaposijevog sarkoma, kriptokoknog meningitisa, pneumocisteze upale pluća. Kliničku sliku bolesti pod utjecajem oportunističke infekcije.
Postupci uzgoja virusa humane imunodeficijencije (HIV)
HIV-1 i HIV-2 mogu se uzgajati u stanicama samo jednog klona TCB4-limfocita - H9, dobivenih iz leukemijskih TCV4-limfocita. U istu svrhu mogu se upotrijebiti i jednoslojne kulture astrocitnih stanica, kod kojih se HIV-1 umnožava. Od životinja do HIV-1 osjetljivih čimpanza.
Otpornost virusa u vanjskom okruženju je niska. Umire pod utjecajem sunčeve svjetlosti i UV zračenja, uništava se na 80 ° C tijekom 30 minuta, kada se liječi uobičajenim sredstvima za dezinfekciju - 20-30 minuta. Za dezinfekciju materijala koji sadrži virus, potrebno je koristiti mikobaktericidne dezinficijense, budući da su učinkoviti protiv mikroorganizama s najvišim otporom.
Laboratorijska dijagnoza HIV infekcije
Glavni način dijagnosticiranja virusa i HIV infekcije je enzimski imunotest. Međutim, s obzirom na činjenicu da ima strukturnu gpl20 i antigene receptore sličnosti nekih ljudskih stanica, uključujući receptore koji obavljaju transport imunoglobulina preko epitelnih stanica sluznice u tijelu mogu pojaviti antitijela vezana antitijela protiv gpl20. U tom slučaju, mogu postojati lažni pozitivni rezultati IFM-a. Stoga, sve pozitivne serume reakcijom proučavali daljnju analizu postupkom iz imunoblot ili Western blot. Metoda se temelji na prepoznavanju testiranih antitijela nakon elektroforetskog razdvajanja i njihovom naknadnom ispitivanje pomoću obilježenih protutijela antispecies. Virološka metoda ima malo koristi zbog složenosti kulture virusa. H9 limfocita klon koristi da se dobije virusni antigeni - nužne komponente dijagnostičkih testovima. CDR metoda omogućuje otkrivanje virusa već u ranoj fazi viremije.
Liječenje infekcije HIV-om
Potrebno je pronaći ili sintetizirati lijekove koji učinkovito inhibiraju aktivnost reverzne transkriptaze (revertaze) ili virusne proteaze. Oni bi spriječili nastanak DNA-provirusa i (ili) inhibiraju intracelularnu multiplicaciju virusa. Suvremena strategija liječenja HIV inficiranih pacijenata koji se temelji na načelu kombiniranu upotrebu lijekova koji inhibiraju virusnu proteazu (jedan od lijekova) i (2 revergazu drugi lijek), - (kombiniranu terapiju trostruku). U Rusiji, za liječenje HIV-inficiranih osoba preporuča kombiniranu uporabu dvaju domaćih lijekova: Crixivan fosfazid i specifično inhibira umnožavanje HIV-a u ranim i kasnim fazama uzgoja, posebno sa smanjenom aktivnošću AZT.
Problem specifične prevencije je potreba za stvaranjem cjepiva koji bi osigurao stvaranje učinkovite imunosti posredovane stanicama na osnovu virusa specifičnih citotoksičnih limfocita bez značajne proizvodnje antitijela. Takav imunitet pružaju Thl-pomoćnici. Moguće je da protutijela, uključujući neutralizaciju virusa, ne samo da su nedjelotvorna u suzbijanju HIV infekcije, već na visokoj razini oni potiskuju imunost posredovanu stanicama. Stoga, anti-HIV cjepivo treba prvenstveno ispuniti dva osnovna zahtjeva: a) biti apsolutno sigurna i b) stimulirati aktivnost T-citotoksičnih limfocita. Proučeno je djelotvornost različitih varijanti cjepiva dobivenih od ubijenih (inaktiviranih) virusa i pojedinačnih antigena s visokim zaštitnim svojstvima. Takvi antigeni mogu biti izolirani iz samih viriona ili kemijski sintetizirani. Predlaže se cjepivo temeljeno na genetskim metodama. To je rekombinantni vaccinia virus koji nosi geni HIV-a koji su odgovorni za sintezu antigena sa snažnim imunogenim svojstvima. Odluka o učinkovitosti ovih cjepiva traje dosta vremena zbog dugog trajanja razdoblja inkubacije HIV infekcije i visoke varijabilnosti patogena. Stvaranje visoko učinkovitog cjepiva protiv HIV-a je hitan temeljni problem.